第四节 体液免疫应答
一、B细胞的活化
根据诱导抗原类型的不同,B细胞可呈现不同的活化方式。
1.TD-Ag诱导的活化自然情况下,多数抗原是TD-Ag,所以B细胞活化多需TH细胞的辅助。B细胞吞噬能力较弱,但其表面Ig是高亲和力的抗原受体,因此可通过受体介导的细胞内摄作用捕获与处理抗原。这个过程需要SIg邻近分子Ig-α和Ig-β的帮助才能传递抗原信号,就象CD3协助TCR一样。抗原刺激构成了B细胞活化的第一信号,可使B细胞初步活化,开始表达粘附分子、MHCⅡ类分子和细胞因子受体等,以便向TH递呈抗原和接受TH细胞的帮助。
B细胞的完全活化产生在抗原递呈的过程中。当TH与B细胞密切接触时,一些对应的协同蛋白可以表现协同刺激作用,例如前述的B7和CD28分子。B细胞表面还有一个特殊的分子CD40,可与TH细胞上的配体CD40L相互作用,所产生的信号足以使B细胞完全活化,甚至可活化未结合抗原的B细胞;这种现象称旁观者效应(bystandereffect)。当CD40L缺乏时,由于B细胞跑得不到TH的帮助而导致一种抗体缺陷病棗超IgM综合征。
B细胞还可通过表面受体(IL-R,FcR,CR等)接受多种因子的作用而促进活化,例如Ils、IgG、C3b和丝裂原等;但是抗原抗体复合物却抑制B细胞的活化。
2.TI-Ag诱导的B细胞活化与TD-Ag不同,TI-Ag(详见第一章)与B细胞上的膜Ig结合时,可通过其大量重复排列的相同表位使B细胞完全活化(图7-4)。但是这种抗原直接的活化作用只能诱导IgM类抗体的产生,而且不能形成记忆细胞,即使多次抗原刺激也不产生再次免疫应答。
3.B细胞的分化与抗体产生B细胞完全活化后,可在淋巴结内,也可迁入骨髓内迅速分化增殖,其中一部分细胞分化为浆细胞。浆细胞是B细胞的终末成熟形式之一,不能继续增殖,而且其寿命仅为数日。但是浆细胞产生抗体的能力特别强,在高峰期一个浆细胞每分钟可分泌数千个抗体分子。一旦抗原刺激解除,抗体应答也会很快消退。
一个增殖克隆的多数B细胞可能不分化成浆细胞,而是返回到静止态变成记忆性B细胞。记忆性B细胞定居于淋巴滤泡内,能存活数年;再被激活时,可重复以前的变化,一部分分化为效应细胞,一部分仍为记忆细胞。数次活化后的子代细胞仍保持原代B细胞的特异性,但中间可能会发生重链的类转换(见第四章)或点突变。这两种变化都不影响B细胞抗原识别的特异性,但点突变影响其产物抗体对抗原的亲和力;高亲和性突变的细胞有生长繁殖的优先权,而低亲和性突变的细胞则选择性死亡。这一现象称为亲和性成熟(affinitymaturation),通过这种机制来保持在后继应答中产生高亲和性的抗体。
二、初次应答与再次应答
1.初次应答抗原初次进入机体后,需首先刺激有限的特异性克隆增殖才能达到足够的反应细胞数,表现为经一定时间的潜伏期才能在血液中检出抗体;如抗原刺激不持续,在应答达到一定之前便消退,多数来不及发生重链的类转换。所以初次应答的显著特点是需要的抗原浓度大、诱导潜伏期长、抗体滴度抵、持续时间短、优势抗体为IgM。
2.再次应答致敏机体受到相同抗原的再次刺激后,在多数情况下会产生再次应答,与初次应答有明显的区别(表7-1)。再次应答可直接活化B记忆细胞,反应性高、增殖快、容易发生类转换;所以表现为潜伏期短、抗体滴度高、持续时间长、优势抗体为IgG和IgA等。免疫应答的这一特性已被广泛应用于传染病的预防、例如疫苗接种一般都做加强免疫,其目就是刺激机体产生再次应答,从而获得对某种传染病更强、更持久的免疫力。
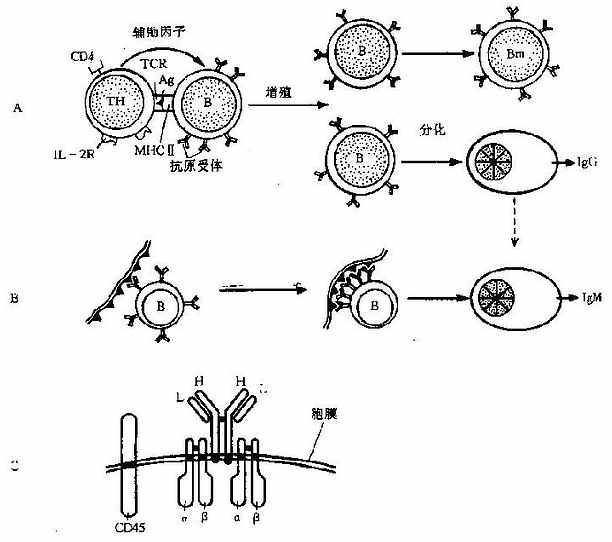
图7-4B细胞的活化
表7-1初次应答与再次应答的比较
初次应答 |
再次应答 | |
抗原递呈 |
非B细胞为主 |
B细胞为主 |
抗原要求 |
较高浓度 |
较低浓度 |
滞后期 |
5~10天 |
2~5天 |
抗体滴度 |
相对低 |
相对高 |
抗体类别 |
IgM为主 |
IgG为主 |
抗原亲和性 |
相对低 |
相对高 |
非特异抗体 |
多见 |
罕见 |
三、抗体的免疫功能
抗体通过与相应抗原的特异性结合发挥生物学效应,最终的生理功能主要是抗感染,其作用机制右归纳为以下几方面:
1.毒素中和作用当抗原为细菌外毒素时,抗原与抗体的结合可中和毒素对宿主的毒性作用。当抗原为激素或酶类时,与抗体结合也可使其活性失活。
2.感染中和作用当病毒与相应抗体结合后,可失去其侵袭细胞的能力,不能进入宿主细胞进行增殖。
3.吞噬调理作用抗菌抗体与细菌结合后,虽不能直接将细胞杀灭,但可作为免疫调理素促进吞噬细胞对细菌的吞噬作用。
4.诱导溶菌作用IgG和IgM类抗体与细菌结合后,可激活补体的经典活化途径裂解病原菌。
5.介导ADCCIgG类抗体还可介导NK细胞等产生ADCC效应,以杀伤病毒感染的靶细胞和恶变细胞。
在另一条件下,抗原抗体反应也可引起机体的组织损伤,例如引起超敏反应或自身免疫病。
, 百拇医药