SARS病毒研究的后基因组时代 SARS研究方向转向分析病毒蛋白及其作用上
美国克罗拉多大学健康科学中心的Hokmes等在Science2003300∶1377上报告,SARS的病原体被确定为SARS病毒(SARS-CoV)后还不到2个月,科学家们已经完成了多株SARS病毒的基因组测序。现在可以对这些来源于不同国家和地区的分离株进行一些比较。
SARS病毒的ORF
SARS病毒基因组具有冠状病毒的所有生物学特征,但与已知冠状病毒存在明显的差异,所以将其归为冠状病毒的一个新的组。多伦多Tor2株和越南Urbani株仅存在8个核苷酸序列的差别,提示在SARS传播过程中,SARS病毒基因组序列保持相对稳定。SARS病毒基因组由5个主要开放读码框(ORF)组成,分别编码RNA聚合酶蛋白、S蛋白、E蛋白、M糖蛋白和N蛋白(图1),其顺序和大小均与其他冠状病毒相似。在主要ORF之间还存在着数量不等的非保守ORF。目前已发现,Tor2株有9个可编码SARS病毒特异性蛋白的潜在ORF,同样,在Urbani株中也发现了其中5个(编码50个氨基酸以上的ORF)。这些蛋白可能不是病毒复制所必需的,也可能在病毒复制和致病过程中发挥特殊功效,还可能起调节宿主对感染的免疫应答的作用。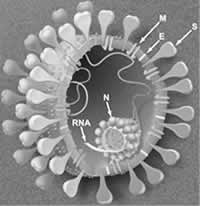
, http://www.100md.com
图1
SARS病毒结构示意图
SARS病毒的TRS
SARS病毒转染细胞的mRNA都具有特征性的3’末端嵌套结构。该结构的5’端都有一个从病毒基因组的5’端衍生出来的长度约70个核苷酸的帽状引导序列。反义RNA病毒的亚基因组mRNA是通过不连续的RNA转录过程合成的,这一过程受靠近每个ORF起始端和引导链3’端的核心转录调控序列(TRS)所调控。Marra等认为SARS病毒TRS的核心序列为5’-CUAAAC-3’,与第2组冠状病毒中鼠和牛冠状病毒的TRS相似;Rota等则认为是5’-AAACGAAC-3’。在已知冠状病毒中,不同基因之间的TRS的差异甚微,而SARS病毒每一个潜在ORF的候选TRS都不一样。SARS病毒的S基因、M基因和ORF10之前都有CUAAAC序列,S基因、M基因、N基因、ORF3、ORF9和ORF10前都有AAACGAAC序列,表明这6个基因在转染细胞中的表达可能是由亚基因组mRNA而来。E基因、ORF7、ORF9和ORF11等上游序列则有显著不同,SARS病毒的E蛋白可能和其他几种冠状病毒的E蛋白一样,是由转录成的一个比较大的mRNA直接翻译而成(而不是亚基因组mRNA)。Rota等用Northern杂交在SARS病毒转染细胞中检出5个高丰量的亚基因组mRNA,而较低丰量的mRNA可能未被检出。需要获得进一步的试验资料,以在转染细胞中确定核心TRS、mRNA的特征和检出编码蛋白。
, 百拇医药
SARS病毒在进化树上的位置
尽管SARS病毒的 3CL蛋白酶(病毒复制酶聚合蛋白的组成部分)、S、E、M和N蛋白的预测氨基酸序列显示,在结构和功能上都与已知冠状病毒的蛋白相似,但其同源蛋白的配对氨基酸序列一致性不及40%~50%。总体而言,SARS病毒基因组和所有已知冠状病毒基因组在进化树上是等距离的。SARS病毒编码聚合酶的基因序列与第2组冠状病毒中牛和鼠冠状病毒最相近,也有某些特征与第3组中的鸟类(冠状)病毒相似。另外,SARS病毒和第3组冠状病毒一样,3’末端都含有由32个核苷酸组成的序列区。与第2组冠状病毒不同,SARS病毒没有编码血凝素的基因。第2组冠状病毒的聚合酶含2个木瓜蛋白样蛋白酶,而SARS病毒和第3组冠状病毒都只编码1个木瓜蛋白样蛋白酶。因此,科学家们都建议,应该将SARS病毒归为冠状病毒的一个新的组。
探寻SARS病毒来源
, http://www.100md.com
很显然,SARS病毒既不是已知冠状病毒的宿主范围突变型,也不是已知冠状病毒之间的简单重组,也不像是基因工程的产物。SARS病毒很可能来源于某个已知冠状病毒的祖先,它在感染人类并产生SARS流行之前,在某种宿主(动物、鸟类或爬行动物)体内经历了一个漫长阶段。确定SARS病毒的来源宿主要依赖于对SARS流行始发地物种的血清学研究,从宿主体内分离出冠状病毒并测定其基因组序列,以确定其为了适应人类宿主所产生的遗传学变化。不同地区的SARS病毒分离株的基因组序列显示了该病毒遗传学的相对稳定性,其序列之间的的小差异可作为流行病学研究的标志物。
SAR病毒后基因组计划
SARS病毒基因组序列的获得,使确定亚基因组mRNA、克隆病毒cDNA、表达重组病毒蛋白并研究这些蛋白在病毒复制和发病机制中的作用成为可能。病毒cDNA和重组病毒蛋白的抗体有助于提高SARS病毒 RNA检测和抗原检测的特异性和敏感性。基因组序列和重组病毒蛋白也有利于促进抗SARS病毒药物和疫苗的研制(图2)。例如,已经用编码SARS病毒 3CL蛋白酶的三维结构模型,来设计可阻断病毒复制的蛋白酶抑制剂。用中和性单克隆抗体被动免疫人体,可达到预防和治疗的目的。通过细胞传代培养,可能可获得减毒活疫苗,然后可由此确定导致病毒毒力下降的突变。对可诱导保护性免疫的抗原的检出,可促进疫苗的研究。目前已经可以通过靶向RNA重组技术和感染性cDNA克隆,以确定决定病毒毒力的抗原决定簇。可表达蛋白、但不具有传染性的基因工程病毒,可成为有效诱导黏膜免疫的疫苗。总而言之,SARS的研究方向已经从最初的病原学研究和病毒基因组测序,转为分析病毒蛋白和这些蛋白在病毒复制和发病机制中的作用,最终目的是获得抗SARS新药和制备疫苗。
图2 SARS病毒的复制过程和药物及疫苗的作用靶位, 百拇医药(于蓝)
SARS病毒的ORF
SARS病毒基因组具有冠状病毒的所有生物学特征,但与已知冠状病毒存在明显的差异,所以将其归为冠状病毒的一个新的组。多伦多Tor2株和越南Urbani株仅存在8个核苷酸序列的差别,提示在SARS传播过程中,SARS病毒基因组序列保持相对稳定。SARS病毒基因组由5个主要开放读码框(ORF)组成,分别编码RNA聚合酶蛋白、S蛋白、E蛋白、M糖蛋白和N蛋白(图1),其顺序和大小均与其他冠状病毒相似。在主要ORF之间还存在着数量不等的非保守ORF。目前已发现,Tor2株有9个可编码SARS病毒特异性蛋白的潜在ORF,同样,在Urbani株中也发现了其中5个(编码50个氨基酸以上的ORF)。这些蛋白可能不是病毒复制所必需的,也可能在病毒复制和致病过程中发挥特殊功效,还可能起调节宿主对感染的免疫应答的作用。
, http://www.100md.com
图1
SARS病毒结构示意图
SARS病毒的TRS
SARS病毒转染细胞的mRNA都具有特征性的3’末端嵌套结构。该结构的5’端都有一个从病毒基因组的5’端衍生出来的长度约70个核苷酸的帽状引导序列。反义RNA病毒的亚基因组mRNA是通过不连续的RNA转录过程合成的,这一过程受靠近每个ORF起始端和引导链3’端的核心转录调控序列(TRS)所调控。Marra等认为SARS病毒TRS的核心序列为5’-CUAAAC-3’,与第2组冠状病毒中鼠和牛冠状病毒的TRS相似;Rota等则认为是5’-AAACGAAC-3’。在已知冠状病毒中,不同基因之间的TRS的差异甚微,而SARS病毒每一个潜在ORF的候选TRS都不一样。SARS病毒的S基因、M基因和ORF10之前都有CUAAAC序列,S基因、M基因、N基因、ORF3、ORF9和ORF10前都有AAACGAAC序列,表明这6个基因在转染细胞中的表达可能是由亚基因组mRNA而来。E基因、ORF7、ORF9和ORF11等上游序列则有显著不同,SARS病毒的E蛋白可能和其他几种冠状病毒的E蛋白一样,是由转录成的一个比较大的mRNA直接翻译而成(而不是亚基因组mRNA)。Rota等用Northern杂交在SARS病毒转染细胞中检出5个高丰量的亚基因组mRNA,而较低丰量的mRNA可能未被检出。需要获得进一步的试验资料,以在转染细胞中确定核心TRS、mRNA的特征和检出编码蛋白。
, 百拇医药
SARS病毒在进化树上的位置
尽管SARS病毒的 3CL蛋白酶(病毒复制酶聚合蛋白的组成部分)、S、E、M和N蛋白的预测氨基酸序列显示,在结构和功能上都与已知冠状病毒的蛋白相似,但其同源蛋白的配对氨基酸序列一致性不及40%~50%。总体而言,SARS病毒基因组和所有已知冠状病毒基因组在进化树上是等距离的。SARS病毒编码聚合酶的基因序列与第2组冠状病毒中牛和鼠冠状病毒最相近,也有某些特征与第3组中的鸟类(冠状)病毒相似。另外,SARS病毒和第3组冠状病毒一样,3’末端都含有由32个核苷酸组成的序列区。与第2组冠状病毒不同,SARS病毒没有编码血凝素的基因。第2组冠状病毒的聚合酶含2个木瓜蛋白样蛋白酶,而SARS病毒和第3组冠状病毒都只编码1个木瓜蛋白样蛋白酶。因此,科学家们都建议,应该将SARS病毒归为冠状病毒的一个新的组。
探寻SARS病毒来源
, http://www.100md.com
很显然,SARS病毒既不是已知冠状病毒的宿主范围突变型,也不是已知冠状病毒之间的简单重组,也不像是基因工程的产物。SARS病毒很可能来源于某个已知冠状病毒的祖先,它在感染人类并产生SARS流行之前,在某种宿主(动物、鸟类或爬行动物)体内经历了一个漫长阶段。确定SARS病毒的来源宿主要依赖于对SARS流行始发地物种的血清学研究,从宿主体内分离出冠状病毒并测定其基因组序列,以确定其为了适应人类宿主所产生的遗传学变化。不同地区的SARS病毒分离株的基因组序列显示了该病毒遗传学的相对稳定性,其序列之间的的小差异可作为流行病学研究的标志物。
SAR病毒后基因组计划
SARS病毒基因组序列的获得,使确定亚基因组mRNA、克隆病毒cDNA、表达重组病毒蛋白并研究这些蛋白在病毒复制和发病机制中的作用成为可能。病毒cDNA和重组病毒蛋白的抗体有助于提高SARS病毒 RNA检测和抗原检测的特异性和敏感性。基因组序列和重组病毒蛋白也有利于促进抗SARS病毒药物和疫苗的研制(图2)。例如,已经用编码SARS病毒 3CL蛋白酶的三维结构模型,来设计可阻断病毒复制的蛋白酶抑制剂。用中和性单克隆抗体被动免疫人体,可达到预防和治疗的目的。通过细胞传代培养,可能可获得减毒活疫苗,然后可由此确定导致病毒毒力下降的突变。对可诱导保护性免疫的抗原的检出,可促进疫苗的研究。目前已经可以通过靶向RNA重组技术和感染性cDNA克隆,以确定决定病毒毒力的抗原决定簇。可表达蛋白、但不具有传染性的基因工程病毒,可成为有效诱导黏膜免疫的疫苗。总而言之,SARS的研究方向已经从最初的病原学研究和病毒基因组测序,转为分析病毒蛋白和这些蛋白在病毒复制和发病机制中的作用,最终目的是获得抗SARS新药和制备疫苗。
图2 SARS病毒的复制过程和药物及疫苗的作用靶位, 百拇医药(于蓝)