生命中的过冷现象
作者:张秋实 张游 李玉明 张诚
单位:张秋实 张游 (承德医学院 医用物理教研室);李玉明 张诚 (低温生理研究室,河北 承德 067000)
关键词:过冷现象;冻结损伤;单细胞;肢体组织;冬眠
中国医学物理学杂志990217 摘要:研究生命中的过冷现象,对于认识其抗寒特性十分有益。本文对于在单细胞深低温保存、高寒地区的动物肢体、动物冬眠时发生的过冷现象及其意义加以介绍。
中图分类号:R381.52
文献标识码:A
文章编号:1005-202X(1999)02-0105-03
前言:在某些情况下,动物组织温度虽然在冰点以下,但不发生冻结,此即过冷现象(Supercooling),但在致冷因素持续作用下降至某温度时过冷状态瓦解迅速发生冻结,该温度即组织的过冷点。水对生命极为重要,脱水、盐化、冻结均可对生命造成不利影响。在高寒地区冻结的影响较为普遍。在有些情况下,由于发生过冷现象抑制冰晶的形成,从而避免了冻结损伤。
, 百拇医药
1 单细胞的过冷与冻结损伤
人类很早就认识到,生物材料能在低温情况下长期保存而不致死亡。但是真正实现生物材料的深低温保存,始于本世纪五十年代。1949年英国剑桥大学的青年学者C*Polge将甘油作为低温保护剂,添加在精子的稀释液中,实现了精子的深低温保存,给生物材料的深低温保存的发展拉开帏幕[1]。此后,生物材料深低温保存技术开始迅速发展。譬如,血细胞的保存和成分输血、造血干细胞的保存和骨髓移植、精子的保存和人工授精、受精卵的保存和胚胎移植,以及角膜、皮肤、骨、细胞株、菌种等的深低温保存均得到广泛的应用。
生物材料虽然能深低温保存,但有时存活率偏低。要提高存活率将涉及许多问题,如冷却与解冻速率,合理的使用低温保护剂及其添加程序等。
实验表明,冷却速率十分重要。不同的生物材料有不同的最佳冷却速度,此速率附近存活率较高,否则存活率下降(图1)。为了解释这种现象,美国学者P*Mazur提出一个理论,认为导致细胞在冷冻过程中死亡的主要因素有两个,一个是细胞膜处于高浓度介质中的时间过长,这在慢速冷却时起主导作用;另一个是细胞内结冰,这在快速冷却时起主导作用。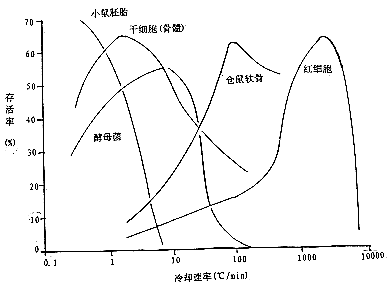
, 百拇医药
图1 表示5种细胞的存活率和冷却速率的关系(仿H.T Meryman and P. Mazur. Crybiology. New Yotk,Academiuc Press,1960)
在现今的低温生物箱中,细胞是置于某种液体介质中冷却。在冷却过程中存在着温度梯度,细胞内液的温度高于细胞外液体介质的温度。当温度降至某一程度时,胞外溶液中一部分水会结晶出来形成纯水的水晶,而剩余的未结冻的液体中溶质浓度将会越来越浓。此时细胞内、外原来的化学平衡被打破,胞内的水分子透过细胞膜向外渗透。在慢速冷却情况下,有足够的时间让水分子从胞内渗透到胞外。如冷却过慢,胞内水分渗出过多造成细胞皱缩,细胞内容质浓度也不断增高,最终细胞内液与胞外介质均变为高浓度溶液。由于电解质可使蛋白质变性,在高浓度溶液作用下细胞膜和细胞器发生损伤,称此种损伤为溶质性损伤。对于溶质性损伤,冷却速率越慢越严重,存活率越低,这是造成细胞死亡的第一个原因。
由于细胞膜对水的渗透率是有限值,在快速冷却时,胞内的水来不及渗 出膜外,细胞外液已经结冰。此时,细胞内液处于过冷状态。过冷状态是一种亚稳定状态,可由于某种条件的变化,使过冷状瓦解,骤然结冰。而冷却速率越快,越容易使细胞内的过冷状态瓦解和发生结冰,这是造成细胞死亡的另一个原因。对于细胞内结冰造成的损伤,冷却速率越慢存活率越高。上述两个因素共同作用使存活率曲线近似抛物线形,如图2所示。
, http://www.100md.com
图2 冷冻过程中高浓度溶质与细胞内结冰共同作用对存活率的影响
(仿H.T Meryman and P.Mazur.Crybiology.New York;Academic Press,1966)
通过上述分析,我们对问题有了定性的认识。若定量的研究这个问题就必须研究水分子透过细胞膜的动力学,建立细胞冻结过程水分子转移的动力学方程。美国学者P.Mazur对此做出贡献并推导出下述方程[2]:
式中V:胞内水体积;T:胞内温度;K:细胞膜对水的通透系数;A:细胞膜有效渗透面积;B:冷却速率;Vw:纯水摩尔体积;P0:细胞外液的水蒸气压;Pi:细胞内液的水蒸气压。
, 百拇医药
该方程是建立在物理化学模型基础上。这里一个重要的假设条件是在冷却过程中细胞内液处于冷状态而不结冰,细胞外介质是在水、冰、溶质共存情况下该方程才能成立。
一般情况,单细胞是在液氮中保存,其温度为-196 ℃。笔者实验发现[3],红细胞、鱼受精卵等生物材料,在添加DMSO、甘油等低温保护剂后以不同降温程序降至-196 ℃时,绝大部分细胞内液发生冻结。虽有少数细胞内液未发生冻结,但在复温解冻时,其过冷的平衡条件遭到破坏,引起细胞内液的重结晶现象。综上所述,细胞深低温保存和解冻过程细胞内液结冰是不可避免的。笔者认为,添加低温保护剂的作用是增加细胞外介质的质点数,降低细胞外介质的冰点和过冷点,尽可能使细胞外介质和细胞内液同时结冰,以避免在冻结过程中过多地引起细胞内失水而发生的溶质性损伤。因此,对细胞冻结损伤的理论应该进一步探讨。
2 温血动物肢体的过冷现象及其耐寒特征
, 百拇医药 温血动物(包括哺乳动物和乌类)能够调节自己的体温,以抵抗环境温度的巨大变化。在高寒地区,譬如黑龙江省北部,冬季野外气温可达-50 ℃,而那里温血动物的体温(Core Temperature)却相当恒定,一般维持在37 ℃附近,冬夏和日夜的变化不超过0.5 ℃。但是,这些动物肢体的组织温度却不是 ,它经常受环境温度的影响,有时很低,甚至在冰点以下处于过冷状态。
1983年在英国剑桥大学召开的第二十届国际低温生物学年会上张诚提出[4],哺乳动物肢体组织的过冷现象是动物为适应寒冷环境而形成的抗寒能力,是进化的产物,是逐渐发展和完善的。这一概念为以后研究动物肢体的抗寒能力冻结损伤开辟了新的途径。笔者实验发现[5-7],凡是能够长时间维持过冷亚稳定状态的肢体一般不出现冻结损伤。但是对于有些肢体,过冷状态一经瓦解随即发生冻结,不论冻结程度深浅复温后均出现不同程度的损伤。所以,组织中冰晶的形成是引起冻结损伤的关键环节。
动物肢体组织的过冷特性与其物理化学性质有关。理论证明[8],过冷点(Tscp)和表面张力(σ)、摩尔质量(M)、冰点(Ti)、密度(ρ)、摩尔潜热(H)及冰胚临界半径(rk)有关,关系式为:Tscp=2σMTi/ρHrk;脂肪的M较大,Ti较低、ρ较小,因此 Tscp较低。所以富含脂肪的组织不仅可阻碍热量散失而且在低温下不易冻结。动物肢体的过冷点还受外环境的影响。笔者实验发现[9],大白鼠尾的过冷点随季节变化,四季中冬至时节过冷点最低,春分最高,夏至与秋分居中。适度的冷习服可增强大鼠的抗寒能力[10],冬季在12 ℃环境温度饲养40天的大鼠,血清中 T3、T4含量升高,鼠尾过冷点降低。但是,在超强寒冷环境-10 ℃饲养大鼠40天后,血清中 T3、T3含量降低,鼠尾过冷点升高。所以,动物在难以耐受的寒冷环境中生活,反而机体的抗寒能力下降。
, http://www.100md.com
笔者实验发现[8,9],兔耳与大白鼠尾的过冷特性与冷却速率有关。冷却速率越慢,过冷点越低,过冷状态越容易发生。空气的密度、比热和热传导系数均较小,因此在寒冷空气中动物肢体容易保持过冷亚稳定状态,不象在液体冷媒中那样容易发生冻结。所以,在高寒地区动物肢体一般不发生冻伤。组织的过冷特性受自身和环境等多因素影响,机理十分复杂,它和冻结损伤有密切关系。
3 动物冬眠与机体整体的过冷现象
冬眠(Hibernation)一词的通常含义是在麻痹或昏睡的状态下越冬[11]。冷血动物的冬眠,哺乳动物和乌类的冬眠,虽然都叫冬眠,但又各不相同。冬季来临,许多地区的气温降至0 ℃以下,大地开始封冻。冷血动物(包括爬行、两栖、鱼类和无脊椎动物)不能调节自己的体温,体温随着环境温度下降,不能维持正常的生命活动,如果不藏起来冬眠,就有被冻死的可能。许多陆生的冷血动物钻入地下冬眠,因为地下比地上温度高,通常在0℃以上,它就不会发生冻结。即使在高寒地区,洞穴温度有时在冰点以下,冷血动物冬眠时也只是浅部组织发生冻结,深部组织和脏器是处于过冷状态[12,13]。由于冬眠时体温很低,能量消耗很少,所以可以完全不吃不喝地度过冬季。如果从地下把它挖出来,就象死的一样,没有明显的感觉表现。当然它没有死,只要把它放在温暖的地方,随着体温的升高会慢慢苏醒过来。但是除非从外界提高冷血动物的体温,它的体温是不会自动提高的。所以,冷血动物的冬眠与苏醒具有被动的特征。
, http://www.100md.com
哺乳类和鸟类的很多目中也有冬眠物种。哺乳类冬眠动物在夏季能维持相当恒定的体温。譬如刺猥(Erinaceus)体温约36 ℃,达乌尔黄鼠(Citellus Dauricus)体温约37 ℃,一般变动范围不超过2 ℃。但在冬季冬眠时则情况大不相同,它们进入凝死的沉睡状态,体温略高于洞穴的温度,华北地区刺猥在冬眠时体温约为2~4 ℃。而在美国阿拉斯加州,北极圈附近冻土带的冬季气候非常恶劣,难以寻觅0 ℃以上的冬眠环境,那里的冬眠哺乳动物经常是全身组织处于过冷状态下冬眠。譬如,北极地鼠(Arctic Ground Squirrel)可在环境温度-20 ℃情况下冬眠,体温(Core Temperature)约为-6 ℃,低于血液冰点温度,即在过冷状态下冬眠[14]。
冷血动物在冬季进入深冬眠后次年春天才醒来。而哺乳类冬眠动物在冬季冬眠时表现出一种周期性的觉醒行为,经过一段时间冬眠后就会自动觉醒。这一周期称为冬眠阵,它包含入眠过程、深眠状态、觉醒过程(出眠)三个时相。在入眠过程体温调定点下移,代谢产热抑制,体温迅速下降,譬如达乌尔黄鼠经过几个小时即从正常体温37 ℃降至2~4 ℃。在深冬眠状态体温最低,此时达乌尔黄鼠体温接近0 ℃;在觉醒过程,体温调定点上移,通过自身产热经过几个小时体温又升高到正常体温37 ℃。不同物种的冬眠动物其冬眠阵不同,少则几天,多则几十天,它们都要经历过若干个冬眠阵才能度过漫长的冬季。特别值得提出的是深冬眠状态的动物可以在更低的环境温度下自动觉醒。譬如将深冬眠的刺猥放在-10 ℃以下的环境,它会自动觉醒,体温迅速上升到正常水平 36 ℃。这是一种本能反应,防止在过于寒冷的情况下被冻死。由此可见,哺乳类冬眠动物不借助外界复温,在很低的环境温度下也能通过自身产热提高体温自动觉醒。所以哺乳动物的冬眠与觉醒具有主动的特征。
, http://www.100md.com
冬眠动物的奇特生理行为一直为低温生物学界所重视。哺乳类动物冬眠是机体整体低温保存的天然模型和理想范例。诱发人类冬眠不仅有重要的理论意义,而且在临床医学、人体整体低温保存和未来的宇宙航行等均有不可估量的应用价值。但是,由于冬眠机理尚不清楚和物种差异,目前诱发人类冬眠还是极其艰巨而又难以实现的事。相信,随着科学的进步和人类对冬眠认识的加深,诱发人类冬眠这一科学构想将来一定会实现。
参考文献
[1] Polge C,et al.Nature,1949,164:666.
[2] Mazur P.The Journal of general physiology,1963,47:347.
[3] 张秋实,等.生理通讯,1997,16(SUPP.1):12.
, http://www.100md.com [4] Zang Cheng,et al.Cryobiology,1983,20:737.
[5] Zhang Cheng,et al.Cryobiology,1985,22:606.
[6] 张 诚,等.应用生理学杂志,1986,2:223.
[7] Zhang Qiushi,et al.Cryobiology,1994,31:577.
[8] 张秋实,等.中国医学物理学杂志,1997,14:140.
[9] 张 诚,等.应用生理学杂志,1985,1:62.
[10] Zhang Cheng,et al.Cryobiology,1987,24:566.
[11] 蔡益鹏主译.哺乳动物和鸟类的冬眠与蛰眠[M].北京:北京大学出版社,1992;1~11.
[12] chmid WD.Cryobiology,1982,19:681.
[13] Zhang Cheng,et al.Cryobiology,1982,19:520.
[14] Barnes BM.Cryobiology,1991,28:551.
收稿日期:1998-05-04, http://www.100md.com
单位:张秋实 张游 (承德医学院 医用物理教研室);李玉明 张诚 (低温生理研究室,河北 承德 067000)
关键词:过冷现象;冻结损伤;单细胞;肢体组织;冬眠
中国医学物理学杂志990217 摘要:研究生命中的过冷现象,对于认识其抗寒特性十分有益。本文对于在单细胞深低温保存、高寒地区的动物肢体、动物冬眠时发生的过冷现象及其意义加以介绍。
中图分类号:R381.52
文献标识码:A
文章编号:1005-202X(1999)02-0105-03
前言:在某些情况下,动物组织温度虽然在冰点以下,但不发生冻结,此即过冷现象(Supercooling),但在致冷因素持续作用下降至某温度时过冷状态瓦解迅速发生冻结,该温度即组织的过冷点。水对生命极为重要,脱水、盐化、冻结均可对生命造成不利影响。在高寒地区冻结的影响较为普遍。在有些情况下,由于发生过冷现象抑制冰晶的形成,从而避免了冻结损伤。
, 百拇医药
1 单细胞的过冷与冻结损伤
人类很早就认识到,生物材料能在低温情况下长期保存而不致死亡。但是真正实现生物材料的深低温保存,始于本世纪五十年代。1949年英国剑桥大学的青年学者C*Polge将甘油作为低温保护剂,添加在精子的稀释液中,实现了精子的深低温保存,给生物材料的深低温保存的发展拉开帏幕[1]。此后,生物材料深低温保存技术开始迅速发展。譬如,血细胞的保存和成分输血、造血干细胞的保存和骨髓移植、精子的保存和人工授精、受精卵的保存和胚胎移植,以及角膜、皮肤、骨、细胞株、菌种等的深低温保存均得到广泛的应用。
生物材料虽然能深低温保存,但有时存活率偏低。要提高存活率将涉及许多问题,如冷却与解冻速率,合理的使用低温保护剂及其添加程序等。
实验表明,冷却速率十分重要。不同的生物材料有不同的最佳冷却速度,此速率附近存活率较高,否则存活率下降(图1)。为了解释这种现象,美国学者P*Mazur提出一个理论,认为导致细胞在冷冻过程中死亡的主要因素有两个,一个是细胞膜处于高浓度介质中的时间过长,这在慢速冷却时起主导作用;另一个是细胞内结冰,这在快速冷却时起主导作用。
, 百拇医药
图1 表示5种细胞的存活率和冷却速率的关系(仿H.T Meryman and P. Mazur. Crybiology. New Yotk,Academiuc Press,1960)
在现今的低温生物箱中,细胞是置于某种液体介质中冷却。在冷却过程中存在着温度梯度,细胞内液的温度高于细胞外液体介质的温度。当温度降至某一程度时,胞外溶液中一部分水会结晶出来形成纯水的水晶,而剩余的未结冻的液体中溶质浓度将会越来越浓。此时细胞内、外原来的化学平衡被打破,胞内的水分子透过细胞膜向外渗透。在慢速冷却情况下,有足够的时间让水分子从胞内渗透到胞外。如冷却过慢,胞内水分渗出过多造成细胞皱缩,细胞内容质浓度也不断增高,最终细胞内液与胞外介质均变为高浓度溶液。由于电解质可使蛋白质变性,在高浓度溶液作用下细胞膜和细胞器发生损伤,称此种损伤为溶质性损伤。对于溶质性损伤,冷却速率越慢越严重,存活率越低,这是造成细胞死亡的第一个原因。
由于细胞膜对水的渗透率是有限值,在快速冷却时,胞内的水来不及渗 出膜外,细胞外液已经结冰。此时,细胞内液处于过冷状态。过冷状态是一种亚稳定状态,可由于某种条件的变化,使过冷状瓦解,骤然结冰。而冷却速率越快,越容易使细胞内的过冷状态瓦解和发生结冰,这是造成细胞死亡的另一个原因。对于细胞内结冰造成的损伤,冷却速率越慢存活率越高。上述两个因素共同作用使存活率曲线近似抛物线形,如图2所示。
, http://www.100md.com
图2 冷冻过程中高浓度溶质与细胞内结冰共同作用对存活率的影响
(仿H.T Meryman and P.Mazur.Crybiology.New York;Academic Press,1966)
通过上述分析,我们对问题有了定性的认识。若定量的研究这个问题就必须研究水分子透过细胞膜的动力学,建立细胞冻结过程水分子转移的动力学方程。美国学者P.Mazur对此做出贡献并推导出下述方程[2]:
式中V:胞内水体积;T:胞内温度;K:细胞膜对水的通透系数;A:细胞膜有效渗透面积;B:冷却速率;Vw:纯水摩尔体积;P0:细胞外液的水蒸气压;Pi:细胞内液的水蒸气压。
, 百拇医药
该方程是建立在物理化学模型基础上。这里一个重要的假设条件是在冷却过程中细胞内液处于冷状态而不结冰,细胞外介质是在水、冰、溶质共存情况下该方程才能成立。
一般情况,单细胞是在液氮中保存,其温度为-196 ℃。笔者实验发现[3],红细胞、鱼受精卵等生物材料,在添加DMSO、甘油等低温保护剂后以不同降温程序降至-196 ℃时,绝大部分细胞内液发生冻结。虽有少数细胞内液未发生冻结,但在复温解冻时,其过冷的平衡条件遭到破坏,引起细胞内液的重结晶现象。综上所述,细胞深低温保存和解冻过程细胞内液结冰是不可避免的。笔者认为,添加低温保护剂的作用是增加细胞外介质的质点数,降低细胞外介质的冰点和过冷点,尽可能使细胞外介质和细胞内液同时结冰,以避免在冻结过程中过多地引起细胞内失水而发生的溶质性损伤。因此,对细胞冻结损伤的理论应该进一步探讨。
2 温血动物肢体的过冷现象及其耐寒特征
, 百拇医药 温血动物(包括哺乳动物和乌类)能够调节自己的体温,以抵抗环境温度的巨大变化。在高寒地区,譬如黑龙江省北部,冬季野外气温可达-50 ℃,而那里温血动物的体温(Core Temperature)却相当恒定,一般维持在37 ℃附近,冬夏和日夜的变化不超过0.5 ℃。但是,这些动物肢体的组织温度却不是 ,它经常受环境温度的影响,有时很低,甚至在冰点以下处于过冷状态。
1983年在英国剑桥大学召开的第二十届国际低温生物学年会上张诚提出[4],哺乳动物肢体组织的过冷现象是动物为适应寒冷环境而形成的抗寒能力,是进化的产物,是逐渐发展和完善的。这一概念为以后研究动物肢体的抗寒能力冻结损伤开辟了新的途径。笔者实验发现[5-7],凡是能够长时间维持过冷亚稳定状态的肢体一般不出现冻结损伤。但是对于有些肢体,过冷状态一经瓦解随即发生冻结,不论冻结程度深浅复温后均出现不同程度的损伤。所以,组织中冰晶的形成是引起冻结损伤的关键环节。
动物肢体组织的过冷特性与其物理化学性质有关。理论证明[8],过冷点(Tscp)和表面张力(σ)、摩尔质量(M)、冰点(Ti)、密度(ρ)、摩尔潜热(H)及冰胚临界半径(rk)有关,关系式为:Tscp=2σMTi/ρHrk;脂肪的M较大,Ti较低、ρ较小,因此 Tscp较低。所以富含脂肪的组织不仅可阻碍热量散失而且在低温下不易冻结。动物肢体的过冷点还受外环境的影响。笔者实验发现[9],大白鼠尾的过冷点随季节变化,四季中冬至时节过冷点最低,春分最高,夏至与秋分居中。适度的冷习服可增强大鼠的抗寒能力[10],冬季在12 ℃环境温度饲养40天的大鼠,血清中 T3、T4含量升高,鼠尾过冷点降低。但是,在超强寒冷环境-10 ℃饲养大鼠40天后,血清中 T3、T3含量降低,鼠尾过冷点升高。所以,动物在难以耐受的寒冷环境中生活,反而机体的抗寒能力下降。
, http://www.100md.com
笔者实验发现[8,9],兔耳与大白鼠尾的过冷特性与冷却速率有关。冷却速率越慢,过冷点越低,过冷状态越容易发生。空气的密度、比热和热传导系数均较小,因此在寒冷空气中动物肢体容易保持过冷亚稳定状态,不象在液体冷媒中那样容易发生冻结。所以,在高寒地区动物肢体一般不发生冻伤。组织的过冷特性受自身和环境等多因素影响,机理十分复杂,它和冻结损伤有密切关系。
3 动物冬眠与机体整体的过冷现象
冬眠(Hibernation)一词的通常含义是在麻痹或昏睡的状态下越冬[11]。冷血动物的冬眠,哺乳动物和乌类的冬眠,虽然都叫冬眠,但又各不相同。冬季来临,许多地区的气温降至0 ℃以下,大地开始封冻。冷血动物(包括爬行、两栖、鱼类和无脊椎动物)不能调节自己的体温,体温随着环境温度下降,不能维持正常的生命活动,如果不藏起来冬眠,就有被冻死的可能。许多陆生的冷血动物钻入地下冬眠,因为地下比地上温度高,通常在0℃以上,它就不会发生冻结。即使在高寒地区,洞穴温度有时在冰点以下,冷血动物冬眠时也只是浅部组织发生冻结,深部组织和脏器是处于过冷状态[12,13]。由于冬眠时体温很低,能量消耗很少,所以可以完全不吃不喝地度过冬季。如果从地下把它挖出来,就象死的一样,没有明显的感觉表现。当然它没有死,只要把它放在温暖的地方,随着体温的升高会慢慢苏醒过来。但是除非从外界提高冷血动物的体温,它的体温是不会自动提高的。所以,冷血动物的冬眠与苏醒具有被动的特征。
, http://www.100md.com
哺乳类和鸟类的很多目中也有冬眠物种。哺乳类冬眠动物在夏季能维持相当恒定的体温。譬如刺猥(Erinaceus)体温约36 ℃,达乌尔黄鼠(Citellus Dauricus)体温约37 ℃,一般变动范围不超过2 ℃。但在冬季冬眠时则情况大不相同,它们进入凝死的沉睡状态,体温略高于洞穴的温度,华北地区刺猥在冬眠时体温约为2~4 ℃。而在美国阿拉斯加州,北极圈附近冻土带的冬季气候非常恶劣,难以寻觅0 ℃以上的冬眠环境,那里的冬眠哺乳动物经常是全身组织处于过冷状态下冬眠。譬如,北极地鼠(Arctic Ground Squirrel)可在环境温度-20 ℃情况下冬眠,体温(Core Temperature)约为-6 ℃,低于血液冰点温度,即在过冷状态下冬眠[14]。
冷血动物在冬季进入深冬眠后次年春天才醒来。而哺乳类冬眠动物在冬季冬眠时表现出一种周期性的觉醒行为,经过一段时间冬眠后就会自动觉醒。这一周期称为冬眠阵,它包含入眠过程、深眠状态、觉醒过程(出眠)三个时相。在入眠过程体温调定点下移,代谢产热抑制,体温迅速下降,譬如达乌尔黄鼠经过几个小时即从正常体温37 ℃降至2~4 ℃。在深冬眠状态体温最低,此时达乌尔黄鼠体温接近0 ℃;在觉醒过程,体温调定点上移,通过自身产热经过几个小时体温又升高到正常体温37 ℃。不同物种的冬眠动物其冬眠阵不同,少则几天,多则几十天,它们都要经历过若干个冬眠阵才能度过漫长的冬季。特别值得提出的是深冬眠状态的动物可以在更低的环境温度下自动觉醒。譬如将深冬眠的刺猥放在-10 ℃以下的环境,它会自动觉醒,体温迅速上升到正常水平 36 ℃。这是一种本能反应,防止在过于寒冷的情况下被冻死。由此可见,哺乳类冬眠动物不借助外界复温,在很低的环境温度下也能通过自身产热提高体温自动觉醒。所以哺乳动物的冬眠与觉醒具有主动的特征。
, http://www.100md.com
冬眠动物的奇特生理行为一直为低温生物学界所重视。哺乳类动物冬眠是机体整体低温保存的天然模型和理想范例。诱发人类冬眠不仅有重要的理论意义,而且在临床医学、人体整体低温保存和未来的宇宙航行等均有不可估量的应用价值。但是,由于冬眠机理尚不清楚和物种差异,目前诱发人类冬眠还是极其艰巨而又难以实现的事。相信,随着科学的进步和人类对冬眠认识的加深,诱发人类冬眠这一科学构想将来一定会实现。
参考文献
[1] Polge C,et al.Nature,1949,164:666.
[2] Mazur P.The Journal of general physiology,1963,47:347.
[3] 张秋实,等.生理通讯,1997,16(SUPP.1):12.
, http://www.100md.com [4] Zang Cheng,et al.Cryobiology,1983,20:737.
[5] Zhang Cheng,et al.Cryobiology,1985,22:606.
[6] 张 诚,等.应用生理学杂志,1986,2:223.
[7] Zhang Qiushi,et al.Cryobiology,1994,31:577.
[8] 张秋实,等.中国医学物理学杂志,1997,14:140.
[9] 张 诚,等.应用生理学杂志,1985,1:62.
[10] Zhang Cheng,et al.Cryobiology,1987,24:566.
[11] 蔡益鹏主译.哺乳动物和鸟类的冬眠与蛰眠[M].北京:北京大学出版社,1992;1~11.
[12] chmid WD.Cryobiology,1982,19:681.
[13] Zhang Cheng,et al.Cryobiology,1982,19:520.
[14] Barnes BM.Cryobiology,1991,28:551.
收稿日期:1998-05-04, http://www.100md.com