视觉电生理(1)
作者:谷万章
单位:哈尔滨242医院眼科150066
关键词:
航空航天医药990337 1 相关的组织解剖及生理学
1.1 细胞内液与细胞外液 这两种液体被细胞膜隔开。细胞膜的通透性比毛细血管壁低,而且有高度的选择性。水、氧和二氧化碳可以自由通过细胞膜,葡萄糖、氨基酸、尿素,各种代谢产物也可以通过细胞膜,而离子却不容易通过细胞膜。
蛋白质和糖原等大分子物质不能通过细胞膜,它们是氨基酸和葡萄糖扩散进入细胞后在细胞内合成的。因而,细胞内液中的蛋白质浓度要比细胞外液中高,一般都在20克%以上。细胞内液的电解质总量多于细胞外液,这主要是由于细胞内液蛋白质含量较高所致,而蛋白质所产生的渗透压却很小,因而细胞内外的渗透压仍是相等的。
, 百拇医药
细胞内液和细胞外液的电解质成分很不相同。细胞外液的正离子以Na+为主,而细胞内液却以K+为主。这不但是由于离子不容易通过细胞膜,而更重要的是,这是细胞膜中的钠泵不断活动的结果。细胞膜对Na+、K+的通透性本来就很低,可是,由于细胞膜内外N a+、K+浓度相差很大,总有一些细胞外液的Na+不断地扩散进入细胞内,又总有一些细胞内液的K+从细胞内扩散到细胞外,钠泵就不断地将细胞内的Na+主动排出到细胞外,同时将细胞外的K+主动转移到细胞内,从而保证了Na+、K+在细胞内、外的不平衡分布。这种不平衡的分布是神经组织和肌肉组织等具有兴奋性的物质基础,也就是当它受到刺激时所发生的细胞生物电现象的来源。细胞内液和细胞外液中的负离子也不相同,细胞外液中的负离子主要是Cl-,而细胞内液中却以有机磷酸离子(HPO2-4)和带负电荷的蛋白质为主,由于后两种负离子不能通过细胞膜且大量存在于细胞内,故细胞内液中的Cl-的浓度就很低了。(见图1)


 图1 各种体液成分的比较
图1 各种体液成分的比较
, 百拇医药
1.2 细胞生物电现象
1.2.1 静息电位 当神经纤维处于静息状态时,细胞膜外是正电位,细胞膜内是负电位,膜内、外有90毫伏的电位差。也就是说,以膜外为零电位,则膜内是-90毫伏。这种电位差叫做静息电位(resting potential)。因而,静息电位就是当神经纤维处于安静状态时存在于细胞膜内外的电位差。这种细胞膜外是正电位,细胞膜内是负电位,二者的电位差稳定于静息电位固定水平的状态,叫做极化(polariged)状态。
静息电位是K+的扩散电位,或者说是K+的电―化学平衡电位。象所有细胞的细胞膜那样,神经纤维的细胞膜对K+和Na+的通透性都很低。可是在静息状态下,相对来说,K +的通透性要比Na+的通透性大20~100倍,所以静息电位是K+的扩散电位。
, 百拇医药
细胞内液中K+的浓度比细胞外液大30倍左右,有很大的浓度梯度,而且细胞膜对K+还有一些通透性,因而就有少量细胞内的K+通过细胞膜扩散到细胞膜外。可是细胞内的负离子大多是大分子的有机磷酸离子和带负电荷的蛋白质,不能通过细胞膜,不能随K+一同扩散出细胞膜而留在细胞之内。这一来,细胞膜外侧有较多的正离子,细胞膜内侧有较多的负离子,因而使膜外电位变得较正,膜内电位变得较负,而使膜内外产生了外正内负的跨膜电位差。这种外正内负的跨膜电位差虽然是由于K+向外扩散而形成,同时这种外正内负的电位差又形成一种阻止正离子K+继续向外扩散的力量,一旦由于浓度梯度而使K+向外扩散的力量和电位差阻止K+向外扩散的力量相等时,也就是两种力量达到平衡时,K+就不再继续向外扩散,或者说K+的净移动等于零,膜两侧的电位差也就稳定于某一固定数值,这就产生了静息电位。所以,静息电位就是K+的扩散电位,或者说是K+的电一化学平衡电位。据计算,胞内的K +只需要扩散出去几百万分之一,即可形成
, 百拇医药
-90毫伏的静息电位。静息电位的高低,是从跨膜电位差大小来考虑的,静息电位降低,叫做除极化,即极化状态减弱;静息电位升高,叫做超极化,即极化状态增强。
1.2.2 动作电位 动作电位需要用电子仪器予以显示,示波器上显示的电位波动,即叫做动作电位(action potential)。动作电位就是正在传导着的兴奋(冲动)所表现的生物电变化。动作电位持续的时间只有0.5~1毫秒。动作电位反映了神经纤维细胞膜内的电位变化,由原来静息状态的-90毫伏,迅速升高到+30毫伏,膜内电位由静息状态的负电位变成了兴奋状态的正电位,出现了极化状态的倒转,即由静息时的外正内负的跨膜电位转变为外负内正,成为倒极化(invertcd potarigation)。膜内电位由-90毫伏上升到0毫伏,为负电位的消失,膜内负电位的消失为除极化,再由0毫伏上升到+30毫伏,称为超射(over shoot)。二者相加,就是动作电位的幅度,一共是120毫伏,这构成了动作电位曲线上升支,也叫做除极化时相(depolarigation phase)。动作电位在神经纤维某一个点上只持续0.5~1 毫秒,因而当上升支迅速到达最高峰后,立即迅速下降到原来的静息电位水平,这就构成了动作电位曲线的下降支,也叫做复极化时相(repolarigation phase)。因而动作电位的曲线很象一个尖锋,所以又可叫做锋电位(splke potential)。
, 百拇医药
动作电位是如何产生的呢?在靠近微电极处用直流电在细胞膜外对神经纤维进行刺激。当通电时,直流电的负极刺激了神经纤维,使受刺激部位膜内外的电位差降低,也就是说发生了除(去)极化。如果直流电太弱,只能引起局部的轻度的除极化,还不足以产生动作电位,因而还不能向外传导。逐渐加大直流电的强度再来刺激,所引起的局部的除极化愈来愈强。一旦除极化达20毫伏左右,即膜内电位降低到-70毫伏左右,就达到了触发动作电位的阈电位(threshold potential)。此时,细胞膜对Na+的通透性突然升高500倍左右,也就是说细胞膜中的钠通道突然大量开放。此时细胞膜对Na+的通透性已超过对K+的通透性。由于细胞外液中Na+浓度比细胞内大10倍左右,而且原来的外正内负的电位差也促使Na+向膜内运动,此时浓度梯度和电位差都促使细胞膜外的一些Na+向细胞膜内扩散,使膜内的正电荷突然暂时增加,使细胞膜暂时处于膜内是正电位、膜外是负电位的倒极化状态,也就形成了动作电位的上升支,即除极化时相。
, 百拇医药
某种强度的刺激恰好足以使细胞膜除极化达到阈电位,从而产生动作电位的最小刺激强度,叫做阈刺激。至于为什么细胞膜的除极化程度达到阈电位时,细胞膜中的钠通道就会突然大量开放,其机制尚不清楚。可以这样合理的推测:细胞膜中的钠通道膜蛋白质是一种贯穿脂质双分子层的膜蛋白质,它平时不开放;一旦跨膜电位改变到了阈电位时,细胞膜上电场力的改变引起钠通道膜蛋白质内部构型发生改变,打开钠通道的“闸门”或者说使之激活(act i vation),使Na+能沿浓度梯度经过钠通道扩散进入胞内。尔后,随着跨膜电位向外负内正方向改变,膜上的电场力又发生改变,使钠通道灭活(inacti vation);加之,钠通道本身有固有的随着时间的延续而灭治的特性,使钠通道在短短的0.5秒左右的时间内又恢复原有构型,重新关闭。
在动作电位的除极化时相到顶点后,立即开始复极化。复极化是K+向膜外扩散所致。钠通道开放的时间只有0.5秒左右,紧接着钠通道的开放,膜的除极也使钾通道开始开放。随着K+通道的开放,在动作电位开始后约0.3 毫秒时,膜对K+的通透性超过了对Na+的通透性,细胞内的K+就由于浓度梯度和倒极化、除极化所形成的电位差,而向细胞外扩散,把正电荷带出细胞膜外,而使跨膜电位又回复到K+的平衡电位,即原来的静息电位水平。这就形成了动作电位的下降支,即复极化时相。总之,动作电位的上升支,即除极化时相,是由于Na+通道开放而使细胞外的Na+扩散进入细胞内而形成的;下降支,即复极化时相,是由于K+通道开放,而使细胞内的K+扩散到细胞外而形成的。这两个过程都是被动的扩散过程,并不消耗能量。
, 百拇医药
细胞内的K+向细胞外扩散,虽然在跨膜电位上来说也是达到了复极化的目的,可是,这样一来,每当神经纤维兴奋一次,细胞内的Na+就稍有增多,K+就稍有减少。当神经纤维多次兴奋之后,细胞内Na+就要逐渐增多,K+就要逐渐减少,势必要改变胞内、外Na+和K+浓度的比值,以致使神经纤维会逐渐失去兴奋的能力。因而还需要复极化的第二个过程,那就是钠泵的运输。细胞内的Na+加多或细胞外的K+加多,都可促进Na+-K+依赖式ATP酶的活动,因而在兴奋之后,钠泵的主动运输加强,将细胞内增多的Na+排出细胞,同时将细胞外的K+运入细胞,终于使细胞内外的Na+和K+的浓度又恢复原状。因而,产生动作电位并不消耗能量,而维持正常的静息电位却要消耗能量,以保证神经纤维的兴奋性能经常保持不变。(见图2、3)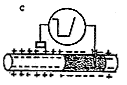
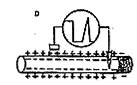
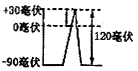
 图2 静息电位与动作电位
图2 静息电位与动作电位
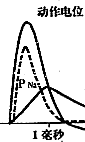
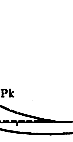
 图3 动作电位及PNa(钠通透性)和PK(钾通透性)的改变
图3 动作电位及PNa(钠通透性)和PK(钾通透性)的改变
, 百拇医药
1.2.3 冲动的传导 冲动不会停留在神经纤维的局部,而要沿着神经纤维向前传导。由于发生动作电位的部位出现了倒极化,跨膜电位是外负内正,而邻接的未兴奋部位仍处于静息状态,跨膜电位是外正内负,而且细胞内液和细胞外液都是导电的,于是就在兴奋部位和邻接的未兴奋部位之间产生局部电流。局部电流流动的方向在细胞外液中是从未兴奋部位流向已兴奋部位,在细胞内液中是从已兴奋部位流向未兴奋部位,其结果就使未兴奋部位膜内电位升高,膜外电位降低,也就是说产生了除极化。未兴奋部位的除极化一旦达到阈电位,细胞膜中的钠通道便突然大量开放,而产生动作电位。冲动就是通过这样的机制迅速向前传导。冲动是一段向前传导着的兴奋波,它的前方不断地向前传导,它的后部又不断地复极化,使兴奋波的长度保持不变。(见图4)
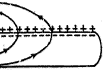 图4 冲动传导时的局部电流
图4 冲动传导时的局部电流
1.2.4 动作电位的触发 把直流电的两个刺激电极放在神经纤维表面,通电时直流电负极刺激了神经纤维,使受刺激部位的跨膜电位差降低,发生了除极化。如果直流电的强度达到了阈刺激,或者超过阈刺激而为阈上刺激,皆可使受刺激部位局部的除极化达到阈电位,从而触发了动作电位。动作电位的波幅大小是由膜内外Na+的浓度梯度和钠通道开放的数目多少决定的,因而动作电位波幅大小并不随阈上刺激的强度大小而有任何改变,因而动作电位是“全或无”式的。动作电位的传导也是通过局部电流引起了邻接尚未兴奋的部位达到阈电位而触发了动作电位,因而,动作电位在神经纤维上的传导是非递减性传导。这就保证了尽管人体有的神经纤维可长达1米以上,并不影响它传导冲动的效率。
, 百拇医药
用直流电负极的阈下刺激来刺激神经纤维,虽不能触发可传导的动作电位向外传导,但并非完全没有作用。阈下刺激可使受到刺激的局部的神经纤维跨膜电位减弱,即出现局部轻度除极化。这既有外来电流的直接作用,也是因为阈下刺激引起局部细胞膜有小量钠通道开放所致。这种局部的低于阈电位的除极化叫做局部阈下反应,简称局部反应。也可叫做局部兴奋状态,简称局部兴奋。局部兴奋不是“全或无”式的,它随刺激强度不同而有低于阈电位的各种不同的强度。局部兴奋虽不能向外传导,但它也可以使邻近的神经纤维跨膜电位出现轻度除极化,这叫做局部兴奋的电紧张性扩布。电紧张性扩布所及范围很小,约为几十几百微米。不但范围很小,而且只要距离稍远一点,除极化程度就急剧减弱,是递减性扩布。这也是局部电流引起的。
局部兴奋的生理意义在于它可以作时间性总和(temporal Sunmation)及空间性总和(Spatial Summation)。先给予一个阈下刺激引起局部兴奋,在它未消失前再给予一个阈下刺激,在原有的局部兴奋基础上再加上一个局部兴奋,就有可能达到阈电位,而触发动作电位向外传导。这就是时间性总和。在邻近的细胞膜同时受到两个或两个以上的阈下刺激时,它们引起的几个局部兴奋也有可能叠加起来达到阈电位,而触发动作电位向外传导,这就是空间性总和。局部兴奋的总和现象在感受器、神经元胞体及骨骼肌运动终板等处十分重要。
, http://www.100md.com
1.3 视网膜的结构与神经递质
1.3.1 视网膜的一般结构 视网膜是一个由三级神经元通过突触联系组成的复杂神经网络,主要包括三层神经细胞三层突触区,并有神经胶质细胞作为支架。
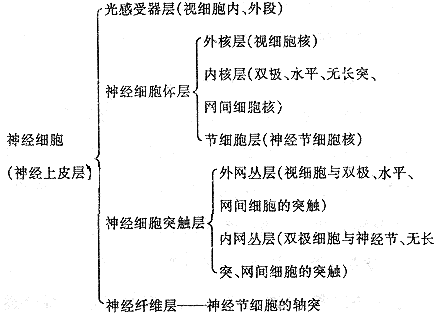
神经胶质细胞――大的Muller神经胶质细胞和其它小神经胶质细胞
1.3.2 视网膜的各类神经细胞 ①视细胞:是光的接收器,高度分化为外段、内段、胞体和末端,按其形态学特征,特别是外段形状的不同,视细胞可分为杆细胞和锥细胞二大类,它们分别主司暗视觉和明视觉、色觉。大多数脊椎动物的锥细胞含有三种不同吸收光谱的视色素,由此命名为红锥、绿锥、兰锥,分别对红、绿、兰光发生反应。视细胞外段由原生质膜内摺成的膜盘堆积组成,在发育过程中,视杆的膜盘除在外段基底部外均与原生质膜分离,而视锥的盘膜绝大部分仍与原生质膜相联。膜蛋白质的主要成分是视色素(视蛋白+祝黄醛),在暗中视色素分子保持一定的空间构型,视细胞的外段原生质膜对Na+通透性较高,有一电流(暗电流)从内段经细胞间隙流向外段,视细胞处于部分去极化状态。光照引起视色素分子构型变化Na+通诱性下降,暗电流减少,细胞处于超极化状态。光信号经过视细胞换能过程后即以电信号形式传向次级神经元。外段盘膜上视色素分子的构型变化如何引起外段质膜上 Na +通透性的改变,曾有种种假说,但目前倾向性的看法是CGMP(环化鸟甘酸)在视细胞内作为细胞内递质(第二信使)起着重要作用。外段膜盘在生命期间不断代谢,放射自显影显示外段基部不断产生新膜盘,顶部不断脱落旧膜盘,其脱落率与自然界的昼和夜以及人为循环光的光照和黑暗期有密切关系,视杆在光照开始后大量脱落膜盘,黑暗时脱落率明显下降,视锥则反之。这种昼夜节律性的“生物钟”现象受到与光有关的内分泌腺和神经活性物质的调控。某些视网膜疾病,如色素变性和夜盲,表现为外段代谢的失调。
, 百拇医药
②双极细胞:相对视细胞,它们是第二级神经元,起着信息传递的作用。双极细胞胞体位于内核层,其树突与视细胞末端小球或小足形成突触(此区构成外网丛层);其轴突末端与神经节细胞的树突或无长突细胞的突起形成突触(此区为内网丛层),从视细胞来的电信号经过双极细胞纵向传至神经节细胞。双极细胞形态多样,分类很多,如按其相接触的视细胞类型和构成的突触类型,可分为陷入型视锥双极细胞、陷入型视杆双极细胞和扁平型视锥双极细胞;若按其对光刺激反应的特征则可分为“撤光型”双极细胞或称“超极化型”双极细胞,和“给光型”双极细胞或称“去极化型”双极细胞。已有工作表明,“超极化型”的轴突分枝终止于内网丛层的a层,而“去极化型”的轴突分枝终止于内网丛层的b层,轴突分枝于a层的是扁平型双极细胞,而于b层的则是陷入型双极细胞。
③水平细胞:位于内核层的外侧,突起广泛分布于外网丛层,在外丛层中起着横向联系和信息调控作用。水平细胞在许多脊椎动物中可分为两大类,一类为短轴突,另一类为无轴突水平细胞。短轴突水平细胞的树突和无轴突水平细胞的突起均与视细胞末端形成带状突触,它们为突触后成分,接受来自视细胞的信息。短轴突水平细胞中的轴突末端与不同神经元形成突触。
, http://www.100md.com
④无长突细胞:是一种无轴突的神经元,胞体位于内核层的内层。按其突起分布特征可分为弥散型无长突细胞(窄弥散型和广弥散型)和分层型无长突细胞(单层、双层和多层型)。若按其化学性能分,目前已知的至少有二十余种。无长突细胞也可分为“撤光型”(其突起分布于内网丛层的a层)、“给光型”(其突起分布于内网丛层的b层)和“撤光-给光型”无长突细胞(其突起分枝终止于a、b二亚层)。
⑤网间细胞:是近十余年来在某些动物中发现的一种新神经元,其胞体位于内核层的无长突细胞之间,从胞体发出的突起伸展于内网丛层和外网丛层二层。在内网丛层突起与无长突细胞形成突触,为突触后成分;而在外网丛层其上升突起与水平细胞或双极细胞突起形成突触,为突触前成分。从内网丛层的无长突细胞来的信息通过网间细胞可以反馈地传向外网丛层的水平细胞,形成了一条离心的反馈通路。
⑥神经节细胞:其胞体组成节细胞层,轴突组成神经纤维层和视神经。视信息在神经节细胞汇总后经视神经传向中枢。神经节细胞按其树突分布特征可分为弥散型和分层型二大类;按其生理特征又可分为“撤光型”(树突分枝于a层)、“给光型”(树突分枝于b层)和“撤光-给光型”(树突分枝于a、b层),以及x、y、w型等。(见图5、6)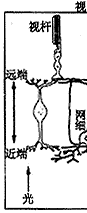
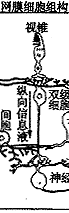
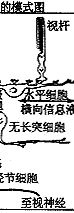
 图5 视网膜细胞组构的模式图
图5 视网膜细胞组构的模式图
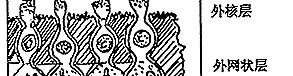
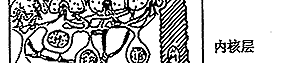
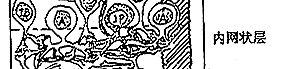
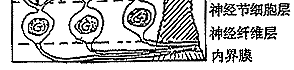 图6 视网膜电子显微镜观察的模式图
图6 视网膜电子显微镜观察的模式图
, 百拇医药
C:视锥细胞 R:视杆细胞 H:水平细胞
IB:陷入型双极细胞,FB:扁平型双极细胞
A:无足细胞,IP:网间细胞,G:神经节细
胞,M:Muller细胞
1.3.3 视网膜神经元间的突触联系和视通路
①突触类型:视网膜的突触基本上分为二大类,化学性突触和电性接触。化学性突触广泛存在于神经元中,是视网膜信息传递的主要突触,包括通常型突触、带状突触和平面接触;电性接触存在于视细胞终末之间、水平细胞之间和无长突细胞之间,称为间隙接触(gopjuncti on)。
②外网丛层:是视细胞轴突与双极细胞、水平细胞或网间细胞突起之间形成的突触区。视细胞末端与水平细胞突起,陷入型双极细胞树突形成带状突触,而与扁平型双极细胞树突形成平面接触。视信息由视细胞传向水平细胞或双极细胞,此外水平细胞与双极细胞之间又形成通常型突触,视信息由水平细胞传至双极细胞。在硬骨鱼中,水平细胞与视细胞形成的带状突触中有特殊的“突触部位”,从水平细胞来的信息可能经此反馈至视细胞。因此,在外网丛层中从视细胞来的信息可以直接通过双极细胞传到神经节细胞,也可通过水平细胞调控后从双极细胞传到节细胞,或从水平细胞反馈至视细胞。
, 百拇医药
③内网丛层:由于包含的神经元多,突触类型和联系方式多而复杂,视信息的传递和调控也极为复杂。尤其是起横向联系的无长突细胞,它的突触类型多种多样,无长突细胞的突起和节细胞的树突在与双极细胞轴突形成的带状突触中为突触后成分,接受双极细胞的视信息;而无长突细胞突起在与双极细胞轴突以及节细胞树突构成的通常型突触中,分别为突触前和突触后,即视信息由无长突细胞传至节细胞或无长突细胞接受来自双极细胞的信息。此外无长突细胞本身之间也有突触联系。无长突细胞突触联系很复杂,有串联式和交叉式等等,在内网丛层中形成了一条局部的神经通路。由此在内网丛层中,视信息可以直接通过双极细胞传到节细胞,也可以通过无长突细胞调控后再传到节细胞。值得注意的是,在某些硬骨鱼中,网间细胞通过其在内网丛层和外网丛层的突触联系,可将视信息从内网丛层反馈至外网丛层;而近期有工作表明一种中枢来的离心纤维与网间细胞的胞体或上升突起形成突触。
综上所述:光信号经过视细胞换能过程中转化为电信号进行传递,传导通路主要是纵向的,视细胞―双极细胞―节细胞;同时还存在着二条横向的调控(水平细胞、无长突细胞);此外,某些动物还有从内网丛层至外网丛层的反馈离心通路和丛中枢来的离心调控通路。
, http://www.100md.com
1.3.9 视网膜的神经递质(NT)和调质(NM) 70年代开始,由于新技术的应用,对视网膜内神经递质、调质的研究取得了显著进展。其中免疫组化方法,不仅能鉴别含NT或NM的神经元,还可追踪其突触联系,从而为了解视信息传递和调控的通路提供了可靠依据。近年来用免疫组化、免疫放射自显影以及生化分析在视网膜中发现多种神经肽或类似的神经活性物质。此外还证明NT和NM可在同一细胞中共存。至今,视网膜中究竟有多少种NT或NM,它们的作用又是什么?仍是当前视觉领域中活跃的研究课题。
目前比较肯定的递质有谷氨酸(GLu)、r-氨基丁酸(GABA)、甘氨酸(GLY)和乙酰胆碱(Acth) ,它们与神经系统中经典的递质一样,从突触前神经细胞中释放后,以扩散形式通过突触间隙,作用于突触后膜受体,改变膜对离子的通透性,快速导致膜电位或膜电阻的变化,使突触后神经元去极化(兴奋性突触)或超极化(抑制性突触)。但一些单胺类如多巴胺(DA),它的作用与众不同,在硬骨鱼视网膜中,网间细胞释放的DA不能直接引起其靶细胞电位或电阻的改变,它的作用是通过细胞内一种酶系统作为第二信使去影响靶细胞。在水平细胞突起表面有一种与环化鸟苷酸(CGMP)有关的DA受体,称受体亚型1(D1),DA作用于D1,胞内C GMP 增高,通过CGMP第二信使作用水平细胞的间隙接触解脱,从而抑制了视信息在水平细胞中的扩散,改变了水平细胞光反应特性。此外在视细胞膜上还有另一种DA受体,称DA受体亚型2( D2),光照时DA释放,通过D2使胞内CGMP水平下降,从而引起了视锥收缩,视杆伸长,以及色素上皮黑色颗粒位移的机械运动。这种通过第二信使起作用的化学物质称之为调质。至于数量甚多的神经肽类物质,它们的作用又是什么?仍需进一步探讨。(待续), 百拇医药
单位:哈尔滨242医院眼科150066
关键词:
航空航天医药990337 1 相关的组织解剖及生理学
1.1 细胞内液与细胞外液 这两种液体被细胞膜隔开。细胞膜的通透性比毛细血管壁低,而且有高度的选择性。水、氧和二氧化碳可以自由通过细胞膜,葡萄糖、氨基酸、尿素,各种代谢产物也可以通过细胞膜,而离子却不容易通过细胞膜。
蛋白质和糖原等大分子物质不能通过细胞膜,它们是氨基酸和葡萄糖扩散进入细胞后在细胞内合成的。因而,细胞内液中的蛋白质浓度要比细胞外液中高,一般都在20克%以上。细胞内液的电解质总量多于细胞外液,这主要是由于细胞内液蛋白质含量较高所致,而蛋白质所产生的渗透压却很小,因而细胞内外的渗透压仍是相等的。
, 百拇医药
细胞内液和细胞外液的电解质成分很不相同。细胞外液的正离子以Na+为主,而细胞内液却以K+为主。这不但是由于离子不容易通过细胞膜,而更重要的是,这是细胞膜中的钠泵不断活动的结果。细胞膜对Na+、K+的通透性本来就很低,可是,由于细胞膜内外N a+、K+浓度相差很大,总有一些细胞外液的Na+不断地扩散进入细胞内,又总有一些细胞内液的K+从细胞内扩散到细胞外,钠泵就不断地将细胞内的Na+主动排出到细胞外,同时将细胞外的K+主动转移到细胞内,从而保证了Na+、K+在细胞内、外的不平衡分布。这种不平衡的分布是神经组织和肌肉组织等具有兴奋性的物质基础,也就是当它受到刺激时所发生的细胞生物电现象的来源。细胞内液和细胞外液中的负离子也不相同,细胞外液中的负离子主要是Cl-,而细胞内液中却以有机磷酸离子(HPO2-4)和带负电荷的蛋白质为主,由于后两种负离子不能通过细胞膜且大量存在于细胞内,故细胞内液中的Cl-的浓度就很低了。(见图1)
图1 各种体液成分的比较, 百拇医药
1.2 细胞生物电现象
1.2.1 静息电位 当神经纤维处于静息状态时,细胞膜外是正电位,细胞膜内是负电位,膜内、外有90毫伏的电位差。也就是说,以膜外为零电位,则膜内是-90毫伏。这种电位差叫做静息电位(resting potential)。因而,静息电位就是当神经纤维处于安静状态时存在于细胞膜内外的电位差。这种细胞膜外是正电位,细胞膜内是负电位,二者的电位差稳定于静息电位固定水平的状态,叫做极化(polariged)状态。
静息电位是K+的扩散电位,或者说是K+的电―化学平衡电位。象所有细胞的细胞膜那样,神经纤维的细胞膜对K+和Na+的通透性都很低。可是在静息状态下,相对来说,K +的通透性要比Na+的通透性大20~100倍,所以静息电位是K+的扩散电位。
, 百拇医药
细胞内液中K+的浓度比细胞外液大30倍左右,有很大的浓度梯度,而且细胞膜对K+还有一些通透性,因而就有少量细胞内的K+通过细胞膜扩散到细胞膜外。可是细胞内的负离子大多是大分子的有机磷酸离子和带负电荷的蛋白质,不能通过细胞膜,不能随K+一同扩散出细胞膜而留在细胞之内。这一来,细胞膜外侧有较多的正离子,细胞膜内侧有较多的负离子,因而使膜外电位变得较正,膜内电位变得较负,而使膜内外产生了外正内负的跨膜电位差。这种外正内负的跨膜电位差虽然是由于K+向外扩散而形成,同时这种外正内负的电位差又形成一种阻止正离子K+继续向外扩散的力量,一旦由于浓度梯度而使K+向外扩散的力量和电位差阻止K+向外扩散的力量相等时,也就是两种力量达到平衡时,K+就不再继续向外扩散,或者说K+的净移动等于零,膜两侧的电位差也就稳定于某一固定数值,这就产生了静息电位。所以,静息电位就是K+的扩散电位,或者说是K+的电一化学平衡电位。据计算,胞内的K +只需要扩散出去几百万分之一,即可形成
, 百拇医药
-90毫伏的静息电位。静息电位的高低,是从跨膜电位差大小来考虑的,静息电位降低,叫做除极化,即极化状态减弱;静息电位升高,叫做超极化,即极化状态增强。
1.2.2 动作电位 动作电位需要用电子仪器予以显示,示波器上显示的电位波动,即叫做动作电位(action potential)。动作电位就是正在传导着的兴奋(冲动)所表现的生物电变化。动作电位持续的时间只有0.5~1毫秒。动作电位反映了神经纤维细胞膜内的电位变化,由原来静息状态的-90毫伏,迅速升高到+30毫伏,膜内电位由静息状态的负电位变成了兴奋状态的正电位,出现了极化状态的倒转,即由静息时的外正内负的跨膜电位转变为外负内正,成为倒极化(invertcd potarigation)。膜内电位由-90毫伏上升到0毫伏,为负电位的消失,膜内负电位的消失为除极化,再由0毫伏上升到+30毫伏,称为超射(over shoot)。二者相加,就是动作电位的幅度,一共是120毫伏,这构成了动作电位曲线上升支,也叫做除极化时相(depolarigation phase)。动作电位在神经纤维某一个点上只持续0.5~1 毫秒,因而当上升支迅速到达最高峰后,立即迅速下降到原来的静息电位水平,这就构成了动作电位曲线的下降支,也叫做复极化时相(repolarigation phase)。因而动作电位的曲线很象一个尖锋,所以又可叫做锋电位(splke potential)。
, 百拇医药
动作电位是如何产生的呢?在靠近微电极处用直流电在细胞膜外对神经纤维进行刺激。当通电时,直流电的负极刺激了神经纤维,使受刺激部位膜内外的电位差降低,也就是说发生了除(去)极化。如果直流电太弱,只能引起局部的轻度的除极化,还不足以产生动作电位,因而还不能向外传导。逐渐加大直流电的强度再来刺激,所引起的局部的除极化愈来愈强。一旦除极化达20毫伏左右,即膜内电位降低到-70毫伏左右,就达到了触发动作电位的阈电位(threshold potential)。此时,细胞膜对Na+的通透性突然升高500倍左右,也就是说细胞膜中的钠通道突然大量开放。此时细胞膜对Na+的通透性已超过对K+的通透性。由于细胞外液中Na+浓度比细胞内大10倍左右,而且原来的外正内负的电位差也促使Na+向膜内运动,此时浓度梯度和电位差都促使细胞膜外的一些Na+向细胞膜内扩散,使膜内的正电荷突然暂时增加,使细胞膜暂时处于膜内是正电位、膜外是负电位的倒极化状态,也就形成了动作电位的上升支,即除极化时相。
, 百拇医药
某种强度的刺激恰好足以使细胞膜除极化达到阈电位,从而产生动作电位的最小刺激强度,叫做阈刺激。至于为什么细胞膜的除极化程度达到阈电位时,细胞膜中的钠通道就会突然大量开放,其机制尚不清楚。可以这样合理的推测:细胞膜中的钠通道膜蛋白质是一种贯穿脂质双分子层的膜蛋白质,它平时不开放;一旦跨膜电位改变到了阈电位时,细胞膜上电场力的改变引起钠通道膜蛋白质内部构型发生改变,打开钠通道的“闸门”或者说使之激活(act i vation),使Na+能沿浓度梯度经过钠通道扩散进入胞内。尔后,随着跨膜电位向外负内正方向改变,膜上的电场力又发生改变,使钠通道灭活(inacti vation);加之,钠通道本身有固有的随着时间的延续而灭治的特性,使钠通道在短短的0.5秒左右的时间内又恢复原有构型,重新关闭。
在动作电位的除极化时相到顶点后,立即开始复极化。复极化是K+向膜外扩散所致。钠通道开放的时间只有0.5秒左右,紧接着钠通道的开放,膜的除极也使钾通道开始开放。随着K+通道的开放,在动作电位开始后约0.3 毫秒时,膜对K+的通透性超过了对Na+的通透性,细胞内的K+就由于浓度梯度和倒极化、除极化所形成的电位差,而向细胞外扩散,把正电荷带出细胞膜外,而使跨膜电位又回复到K+的平衡电位,即原来的静息电位水平。这就形成了动作电位的下降支,即复极化时相。总之,动作电位的上升支,即除极化时相,是由于Na+通道开放而使细胞外的Na+扩散进入细胞内而形成的;下降支,即复极化时相,是由于K+通道开放,而使细胞内的K+扩散到细胞外而形成的。这两个过程都是被动的扩散过程,并不消耗能量。
, 百拇医药
细胞内的K+向细胞外扩散,虽然在跨膜电位上来说也是达到了复极化的目的,可是,这样一来,每当神经纤维兴奋一次,细胞内的Na+就稍有增多,K+就稍有减少。当神经纤维多次兴奋之后,细胞内Na+就要逐渐增多,K+就要逐渐减少,势必要改变胞内、外Na+和K+浓度的比值,以致使神经纤维会逐渐失去兴奋的能力。因而还需要复极化的第二个过程,那就是钠泵的运输。细胞内的Na+加多或细胞外的K+加多,都可促进Na+-K+依赖式ATP酶的活动,因而在兴奋之后,钠泵的主动运输加强,将细胞内增多的Na+排出细胞,同时将细胞外的K+运入细胞,终于使细胞内外的Na+和K+的浓度又恢复原状。因而,产生动作电位并不消耗能量,而维持正常的静息电位却要消耗能量,以保证神经纤维的兴奋性能经常保持不变。(见图2、3)
图2 静息电位与动作电位图3 动作电位及PNa(钠通透性)和PK(钾通透性)的改变, 百拇医药
1.2.3 冲动的传导 冲动不会停留在神经纤维的局部,而要沿着神经纤维向前传导。由于发生动作电位的部位出现了倒极化,跨膜电位是外负内正,而邻接的未兴奋部位仍处于静息状态,跨膜电位是外正内负,而且细胞内液和细胞外液都是导电的,于是就在兴奋部位和邻接的未兴奋部位之间产生局部电流。局部电流流动的方向在细胞外液中是从未兴奋部位流向已兴奋部位,在细胞内液中是从已兴奋部位流向未兴奋部位,其结果就使未兴奋部位膜内电位升高,膜外电位降低,也就是说产生了除极化。未兴奋部位的除极化一旦达到阈电位,细胞膜中的钠通道便突然大量开放,而产生动作电位。冲动就是通过这样的机制迅速向前传导。冲动是一段向前传导着的兴奋波,它的前方不断地向前传导,它的后部又不断地复极化,使兴奋波的长度保持不变。(见图4)
图4 冲动传导时的局部电流1.2.4 动作电位的触发 把直流电的两个刺激电极放在神经纤维表面,通电时直流电负极刺激了神经纤维,使受刺激部位的跨膜电位差降低,发生了除极化。如果直流电的强度达到了阈刺激,或者超过阈刺激而为阈上刺激,皆可使受刺激部位局部的除极化达到阈电位,从而触发了动作电位。动作电位的波幅大小是由膜内外Na+的浓度梯度和钠通道开放的数目多少决定的,因而动作电位波幅大小并不随阈上刺激的强度大小而有任何改变,因而动作电位是“全或无”式的。动作电位的传导也是通过局部电流引起了邻接尚未兴奋的部位达到阈电位而触发了动作电位,因而,动作电位在神经纤维上的传导是非递减性传导。这就保证了尽管人体有的神经纤维可长达1米以上,并不影响它传导冲动的效率。
, 百拇医药
用直流电负极的阈下刺激来刺激神经纤维,虽不能触发可传导的动作电位向外传导,但并非完全没有作用。阈下刺激可使受到刺激的局部的神经纤维跨膜电位减弱,即出现局部轻度除极化。这既有外来电流的直接作用,也是因为阈下刺激引起局部细胞膜有小量钠通道开放所致。这种局部的低于阈电位的除极化叫做局部阈下反应,简称局部反应。也可叫做局部兴奋状态,简称局部兴奋。局部兴奋不是“全或无”式的,它随刺激强度不同而有低于阈电位的各种不同的强度。局部兴奋虽不能向外传导,但它也可以使邻近的神经纤维跨膜电位出现轻度除极化,这叫做局部兴奋的电紧张性扩布。电紧张性扩布所及范围很小,约为几十几百微米。不但范围很小,而且只要距离稍远一点,除极化程度就急剧减弱,是递减性扩布。这也是局部电流引起的。
局部兴奋的生理意义在于它可以作时间性总和(temporal Sunmation)及空间性总和(Spatial Summation)。先给予一个阈下刺激引起局部兴奋,在它未消失前再给予一个阈下刺激,在原有的局部兴奋基础上再加上一个局部兴奋,就有可能达到阈电位,而触发动作电位向外传导。这就是时间性总和。在邻近的细胞膜同时受到两个或两个以上的阈下刺激时,它们引起的几个局部兴奋也有可能叠加起来达到阈电位,而触发动作电位向外传导,这就是空间性总和。局部兴奋的总和现象在感受器、神经元胞体及骨骼肌运动终板等处十分重要。
, http://www.100md.com
1.3 视网膜的结构与神经递质
1.3.1 视网膜的一般结构 视网膜是一个由三级神经元通过突触联系组成的复杂神经网络,主要包括三层神经细胞三层突触区,并有神经胶质细胞作为支架。
神经胶质细胞――大的Muller神经胶质细胞和其它小神经胶质细胞
1.3.2 视网膜的各类神经细胞 ①视细胞:是光的接收器,高度分化为外段、内段、胞体和末端,按其形态学特征,特别是外段形状的不同,视细胞可分为杆细胞和锥细胞二大类,它们分别主司暗视觉和明视觉、色觉。大多数脊椎动物的锥细胞含有三种不同吸收光谱的视色素,由此命名为红锥、绿锥、兰锥,分别对红、绿、兰光发生反应。视细胞外段由原生质膜内摺成的膜盘堆积组成,在发育过程中,视杆的膜盘除在外段基底部外均与原生质膜分离,而视锥的盘膜绝大部分仍与原生质膜相联。膜蛋白质的主要成分是视色素(视蛋白+祝黄醛),在暗中视色素分子保持一定的空间构型,视细胞的外段原生质膜对Na+通透性较高,有一电流(暗电流)从内段经细胞间隙流向外段,视细胞处于部分去极化状态。光照引起视色素分子构型变化Na+通诱性下降,暗电流减少,细胞处于超极化状态。光信号经过视细胞换能过程后即以电信号形式传向次级神经元。外段盘膜上视色素分子的构型变化如何引起外段质膜上 Na +通透性的改变,曾有种种假说,但目前倾向性的看法是CGMP(环化鸟甘酸)在视细胞内作为细胞内递质(第二信使)起着重要作用。外段膜盘在生命期间不断代谢,放射自显影显示外段基部不断产生新膜盘,顶部不断脱落旧膜盘,其脱落率与自然界的昼和夜以及人为循环光的光照和黑暗期有密切关系,视杆在光照开始后大量脱落膜盘,黑暗时脱落率明显下降,视锥则反之。这种昼夜节律性的“生物钟”现象受到与光有关的内分泌腺和神经活性物质的调控。某些视网膜疾病,如色素变性和夜盲,表现为外段代谢的失调。
, 百拇医药
②双极细胞:相对视细胞,它们是第二级神经元,起着信息传递的作用。双极细胞胞体位于内核层,其树突与视细胞末端小球或小足形成突触(此区构成外网丛层);其轴突末端与神经节细胞的树突或无长突细胞的突起形成突触(此区为内网丛层),从视细胞来的电信号经过双极细胞纵向传至神经节细胞。双极细胞形态多样,分类很多,如按其相接触的视细胞类型和构成的突触类型,可分为陷入型视锥双极细胞、陷入型视杆双极细胞和扁平型视锥双极细胞;若按其对光刺激反应的特征则可分为“撤光型”双极细胞或称“超极化型”双极细胞,和“给光型”双极细胞或称“去极化型”双极细胞。已有工作表明,“超极化型”的轴突分枝终止于内网丛层的a层,而“去极化型”的轴突分枝终止于内网丛层的b层,轴突分枝于a层的是扁平型双极细胞,而于b层的则是陷入型双极细胞。
③水平细胞:位于内核层的外侧,突起广泛分布于外网丛层,在外丛层中起着横向联系和信息调控作用。水平细胞在许多脊椎动物中可分为两大类,一类为短轴突,另一类为无轴突水平细胞。短轴突水平细胞的树突和无轴突水平细胞的突起均与视细胞末端形成带状突触,它们为突触后成分,接受来自视细胞的信息。短轴突水平细胞中的轴突末端与不同神经元形成突触。
, http://www.100md.com
④无长突细胞:是一种无轴突的神经元,胞体位于内核层的内层。按其突起分布特征可分为弥散型无长突细胞(窄弥散型和广弥散型)和分层型无长突细胞(单层、双层和多层型)。若按其化学性能分,目前已知的至少有二十余种。无长突细胞也可分为“撤光型”(其突起分布于内网丛层的a层)、“给光型”(其突起分布于内网丛层的b层)和“撤光-给光型”无长突细胞(其突起分枝终止于a、b二亚层)。
⑤网间细胞:是近十余年来在某些动物中发现的一种新神经元,其胞体位于内核层的无长突细胞之间,从胞体发出的突起伸展于内网丛层和外网丛层二层。在内网丛层突起与无长突细胞形成突触,为突触后成分;而在外网丛层其上升突起与水平细胞或双极细胞突起形成突触,为突触前成分。从内网丛层的无长突细胞来的信息通过网间细胞可以反馈地传向外网丛层的水平细胞,形成了一条离心的反馈通路。
⑥神经节细胞:其胞体组成节细胞层,轴突组成神经纤维层和视神经。视信息在神经节细胞汇总后经视神经传向中枢。神经节细胞按其树突分布特征可分为弥散型和分层型二大类;按其生理特征又可分为“撤光型”(树突分枝于a层)、“给光型”(树突分枝于b层)和“撤光-给光型”(树突分枝于a、b层),以及x、y、w型等。(见图5、6)
图5 视网膜细胞组构的模式图图6 视网膜电子显微镜观察的模式图, 百拇医药
C:视锥细胞 R:视杆细胞 H:水平细胞
IB:陷入型双极细胞,FB:扁平型双极细胞
A:无足细胞,IP:网间细胞,G:神经节细
胞,M:Muller细胞
1.3.3 视网膜神经元间的突触联系和视通路
①突触类型:视网膜的突触基本上分为二大类,化学性突触和电性接触。化学性突触广泛存在于神经元中,是视网膜信息传递的主要突触,包括通常型突触、带状突触和平面接触;电性接触存在于视细胞终末之间、水平细胞之间和无长突细胞之间,称为间隙接触(gopjuncti on)。
②外网丛层:是视细胞轴突与双极细胞、水平细胞或网间细胞突起之间形成的突触区。视细胞末端与水平细胞突起,陷入型双极细胞树突形成带状突触,而与扁平型双极细胞树突形成平面接触。视信息由视细胞传向水平细胞或双极细胞,此外水平细胞与双极细胞之间又形成通常型突触,视信息由水平细胞传至双极细胞。在硬骨鱼中,水平细胞与视细胞形成的带状突触中有特殊的“突触部位”,从水平细胞来的信息可能经此反馈至视细胞。因此,在外网丛层中从视细胞来的信息可以直接通过双极细胞传到神经节细胞,也可通过水平细胞调控后从双极细胞传到节细胞,或从水平细胞反馈至视细胞。
, 百拇医药
③内网丛层:由于包含的神经元多,突触类型和联系方式多而复杂,视信息的传递和调控也极为复杂。尤其是起横向联系的无长突细胞,它的突触类型多种多样,无长突细胞的突起和节细胞的树突在与双极细胞轴突形成的带状突触中为突触后成分,接受双极细胞的视信息;而无长突细胞突起在与双极细胞轴突以及节细胞树突构成的通常型突触中,分别为突触前和突触后,即视信息由无长突细胞传至节细胞或无长突细胞接受来自双极细胞的信息。此外无长突细胞本身之间也有突触联系。无长突细胞突触联系很复杂,有串联式和交叉式等等,在内网丛层中形成了一条局部的神经通路。由此在内网丛层中,视信息可以直接通过双极细胞传到节细胞,也可以通过无长突细胞调控后再传到节细胞。值得注意的是,在某些硬骨鱼中,网间细胞通过其在内网丛层和外网丛层的突触联系,可将视信息从内网丛层反馈至外网丛层;而近期有工作表明一种中枢来的离心纤维与网间细胞的胞体或上升突起形成突触。
综上所述:光信号经过视细胞换能过程中转化为电信号进行传递,传导通路主要是纵向的,视细胞―双极细胞―节细胞;同时还存在着二条横向的调控(水平细胞、无长突细胞);此外,某些动物还有从内网丛层至外网丛层的反馈离心通路和丛中枢来的离心调控通路。
, http://www.100md.com
1.3.9 视网膜的神经递质(NT)和调质(NM) 70年代开始,由于新技术的应用,对视网膜内神经递质、调质的研究取得了显著进展。其中免疫组化方法,不仅能鉴别含NT或NM的神经元,还可追踪其突触联系,从而为了解视信息传递和调控的通路提供了可靠依据。近年来用免疫组化、免疫放射自显影以及生化分析在视网膜中发现多种神经肽或类似的神经活性物质。此外还证明NT和NM可在同一细胞中共存。至今,视网膜中究竟有多少种NT或NM,它们的作用又是什么?仍是当前视觉领域中活跃的研究课题。
目前比较肯定的递质有谷氨酸(GLu)、r-氨基丁酸(GABA)、甘氨酸(GLY)和乙酰胆碱(Acth) ,它们与神经系统中经典的递质一样,从突触前神经细胞中释放后,以扩散形式通过突触间隙,作用于突触后膜受体,改变膜对离子的通透性,快速导致膜电位或膜电阻的变化,使突触后神经元去极化(兴奋性突触)或超极化(抑制性突触)。但一些单胺类如多巴胺(DA),它的作用与众不同,在硬骨鱼视网膜中,网间细胞释放的DA不能直接引起其靶细胞电位或电阻的改变,它的作用是通过细胞内一种酶系统作为第二信使去影响靶细胞。在水平细胞突起表面有一种与环化鸟苷酸(CGMP)有关的DA受体,称受体亚型1(D1),DA作用于D1,胞内C GMP 增高,通过CGMP第二信使作用水平细胞的间隙接触解脱,从而抑制了视信息在水平细胞中的扩散,改变了水平细胞光反应特性。此外在视细胞膜上还有另一种DA受体,称DA受体亚型2( D2),光照时DA释放,通过D2使胞内CGMP水平下降,从而引起了视锥收缩,视杆伸长,以及色素上皮黑色颗粒位移的机械运动。这种通过第二信使起作用的化学物质称之为调质。至于数量甚多的神经肽类物质,它们的作用又是什么?仍需进一步探讨。(待续), 百拇医药