神经酰胺与胞内信号传递
作者:姚毓奇
单位:中山医科大学肿瘤研究所治疗基础室,(广州,510060)
关键词:信号传递;神经酰胺;生物学意义
癌症990539
曾益新(审校)
中图分类号:R-05;R329.28;R344 文献标识码:A
文章编号:1000-467X(1999)05-0605-03
(神经)鞘磷脂(Sphingomyelin,SM)长期以来被认为只是生物膜的一个结构成分,主要分布在脂双层的外层,不知它有什么功能。然而,自从1991年Kolesnick等首先报道甘油二酯(DAG)可诱导SM水解生成神经酰胺(Ceramide,Cer),并发现Cer本身不能象DAG那样激活PKC以来,Cer作为胞内第二信使得到了广泛的证实。作为第二信使,Cer在介导细胞生长、增殖、分化、凋亡及损伤等多种生理、病理过程中起着广泛的作用。本文就Cer生成相关的酶、Cer的效应分子及其与糖脂和其它鞘脂代谢物[如鞘氨醇(Sphingosine)、鞘氨醇1磷酸(Sphingosine-1-Phospha-te)]介导的信号传递途径之间的联系作一综述,并简述Cer信使途径的生物学意义。
, 百拇医药
1 参与Cer生成的酶
1.1 鞘磷脂酶(Sphingomyelinas-e,SMase)
SM可在SMase催化下水解产生Cer和磷酸胆碱,此过程是可逆的,因此称为SM循环。多种激活因素(如TNF-α、IFN-γ、ILs、FasL、1,25-(OH)2-D3、地塞米松、化疗药物、电离辐射及热刺激等)可诱导诸如U937、HL-60、JurkatT细胞、Molt4、PC12等细胞内SM水解而产生Cer,最终导致细胞周期停滞或凋亡等现象的发生。根据SMase的体外活化条件及最适pH值的不同,现发现至少存在4种SMase:(1)膜型中性SMase(nSMase),其活性依赖于Mg2+及中性pH环境(最适pH7.0~7.5),(2)胞浆型nSMase,为Mg2+非依赖性。BinLiu等报道该酶可被生理浓度的GSH抑制,GSH合成抑制剂可导致时间依赖性的GSH降低,而在许多细胞损伤和凋亡过程中发现有GSH的减少,提示GSH的降低可能是nSMase活化的重要机制。(3)酸性SMase(aSMase),主要分布于溶酶体及核内体,激活时需DAG,最适pH值4.5~5.0。(4)碱性SMase,激活时需胆盐,最适pH值9.0。前三种参与信号传递已得到确认,碱性SMase是否与信号传递相关有待进一步研究证实,其中nSMase被认为是SM循环的调节性激活和信号传递中的关键性酶。人的nSMase和aSMase已被克隆,其中nSMase为47.6KD的糖蛋白,由423个氨基酸残基组成。至今测得的具有最强的nSMase活性的组织是中枢神经系统,这与nSMase的广泛组织分布不相一致,提示可能存在一个脑组织特异性的nSMase的活化过程〔1~3〕。
, 百拇医药
1.2 Cer酶(Ceramidase)和Cer合酶(Ceramide Synthase)
Cer在Ceramidase催化下脱酰基生成鞘氨醇,鞘氨醇也可在Cer合酶催化下生成Cer,因此这二种酶在控制细胞内Cer水平起重要作用。Cer可通过激活Cer合酶而从头合成,但至今未发现鞘氨醇头合成途径,因此体内鞘氨醇的生成必需依赖于Ceramidase。许多实验发现Cer和鞘氨醇常诱导相反或不同的生物效应,因此这两种可相互转变的鞘脂代谢物生成的平衡对调控细胞功能可能是一个重要因素。此外,发现诱导细胞凋亡发生的Cer不一定只来自SM的水解:抗癌药物daunorubicin可刺激P388和U937细胞内Cer水平升高并发生凋亡,但此种Cer是通过激活Cer合酶而从头合成的,这是从研究fumonisins得到的结果。Fumonisins是一类Cer合酶的天然的、特异性抑制物,FumonisinB1(FB1)可抑制双氢鞘氨醇-N-酰基转移酶(Sphinganine N-acyltransferase),即Cer合酶的活性而阻断daunorubicin引发的Cer升高及凋亡。Garzotto等报道TPA通过激活PKC-α而诱导LNCaP前列腺细胞凋亡是由Cer合酶介导的,因为此过程中aSMase和nSMase活性均未见升高,并且FB1可取消TPA诱导的Cer产生和细胞凋亡〔1,4〕。
, 百拇医药
1.3 磷脂酸磷酸水解酶(phosphatidate phohydrolase,PAP)
PAP以两种形式存在,PAP-1存在于胞浆,为Mg2+依赖性,可转移至内质网或线粒体,也可转移至细胞膜内表面,并降解由PLD活化产生的磷脂酸(PA)而生成DAG,但PAP-1是否参与信号传递有待证实〔1〕。PAP-2是一膜结合蛋白,为Mg2+非依赖性,也可降解PA产生DAG,但这种PLD来源的DAG不能激活PKC,因此PAP-2可能是通过PA而终止信号传递。PAP-2也可降解Cer-1-P和Sph-1-P(SPP),因此PAP-2可通过降解这类物质而终止来自PA、Cer-1-P、Sph-10-P的信号,但也会产生DAG、Cer和Spingosine(SP)一类具有生物活性的物质,可见PAP可通过调节这类参与细胞活化和控制细胞生长的生物活性脂质之间的平衡而在细胞信号传递中起重要作用〔1〕。
, http://www.100md.com
2 参与Cer信号传递的效应分子
Cer作为第二信使,必定要通过一系列效应分子的级联放大反应,最终激活核内相关基因而产生不同的生物学效应。目前发现神经酰胺活化的蛋白磷酸酶(Ceramide-activated protein phosphatase,CAPP)、神经酰胺活化的蛋白激酶(Ceramide-activated profein kinase,CAPK)、PKC-ζ及原癌基因产物VaV等是Cer的直接靶分子〔5〕。此外,最近在不同细胞系中还发现有下列几种效应分子参与Cer介导的信号传递。
2.1 JNK和ERK
JNK(C-Jun N-terminal Kinase),又称SAPK(应激活化的蛋白激酶),是MAPK超家族的亚族成员,JNK介导的信号传递通路与细胞生长抑制、凋亡及炎症反应相关;ERK(extracellular signal-regulated kinase)途径主要与细胞增殖相关,生长因子和鞘氨醇等可激活ERK而促进细胞生长。实验资料表明,Cer可迅速激活P46-JNK1/P54-JNK2,从而增强JNK的主要底物C-Jun的表达和活性增高,同时Cer可使P42-ERK1/P44-ERK2活性逐渐地、中等程度地降低,而且发现JNK的底物C-Jun的表达和活性的增强对Cer诱导的细胞凋亡发生是绝对必需的〔6〕。但有人在以C2-Cer处理含有Ki-ras突变的人HCT-116结肠癌细胞研究中发现,尽管C2-Cer引起该细胞显著凋亡,但没有观察到JNK/SAPK的激活,可能在该类细胞中有其他信号途径参与了Cer诱导的凋亡〔7〕。
, 百拇医药
2.2 TAK1
TAK1(TGF-βactivated kinase),为丝氨酸/苏氨酸蛋白激酶,属于MAPKKK家族成员,人和鼠的TAK1氨基酸序列高度同源(同源性达98%),因此TAK1是进化上相当保守的激酶。据报道,Cer可直接激活TAK1,活化的TAK1导致SEK1/MKK4激活,后者可继续活化JNK/SAPK,而TAK1的非活化型阻止Cer诱导的JNK/SAPK的激活,说明TAK1是Cer活化JNK/SAPK的关键性中间物,其它脂类物质(包括DAG,花生四烯酸)处理细胞时发现TAK1未被激活,提示TAK1参与了Cer诱发的信号传递〔8〕。此外,还发现TAK1结合蛋白(TAB1)的过表达几乎完全抑制TGF-β刺激TAK1的活化,但对Cer诱导的TAK1激活作用甚微,因此Cer对TAK1的激活是TAB1非依赖性的〔11〕。
2.3 Caspases
, http://www.100md.com
Caspases,属于天冬氨酸特异的半胱氨酸蛋白酶家族,目前根据其种系发生可分为三个亚族:(1)CED-3亚族,包括Caspase-3/CPP32、Caspase-6、Caspase-7、Caspase-8、Caspase-9和Caspase-10;(2)ICE家族,包括Caspase-1、Caspase-4、Caspase-5、Caspase-11;(3)Nedd-2亚族,以Caspase-2为代表。DNA修复酶PARP、DNA-PKcs和核结构蛋白NuMa、lamins等是这类酶的底物,这些底物的水解可能与细胞凋亡时核形态和生化上的改变有关。CED-3亚族的几个Caspase可在凋亡早期催化PARP的分解,其中Caspase-3/CPP32被认为最有效(其次为Caspase-7);核lamins的降解是浓缩染色质进入凋亡小体(凋亡晚期典型的形态结构)所必需的,Caspase-6是目前唯一所知的水解lamins的酶〔10,11〕。Kuo等报道Caspase-3在C2-Cer诱导的HL-60细胞凋亡过程中被激活,而Caspase-3的特异性抑制物Ac-DEVD-CHO可有效地保护Cer诱导的细胞死亡,提示Caspase-3在Cer诱导的凋亡过程中起关键作用〔12〕
, http://www.100md.com
3 Cer信使途径与糖脂及其他鞘脂代谢物介导的信号途径之间的相互作用
许多文献报道了Cer能够诱导细胞凋亡而DAG具有抗凋亡的特性,并且在多种类型细胞中发现因Cer水平增高而诱导的细胞死亡可被DAG所抑制,因此在糖脂和鞘脂信号途经之间必然存在相互作用的位点。目前发现以下事实:(1)细胞渗透性C2和C6-Cer可抑制诸如血清、凝血酶、磷脂酸、SPP或佛波酯等PLD(磷脂酶D)激活剂刺激的PLD活性的增加,而双氢Cer对此无多大影响,说明Cer作用具有特异性;(2)PKC是一类由多种同功酶组成的结构相关的丝氨酸/苏氨酸蛋白酶激酶,与许多细胞功能相关。目前只发现PKC-α、δ、ε受Cer调节。最近有报道称Cer可诱导PKC-δ、ε从胞膜向胞浆的转位,并且外源SMase、TNF-α、Fas抗体(均可诱导凋亡发生)有类似功能,提示PKC-δ、ε的转位在Cer介导的凋亡中起重要作用。(3)细胞渗透性Cer抑制PA、lysoPA、SPP、C2-/C8-Cer-1-P及天然Cer-1-P刺激的DNA合成,即抑制了这类物质对细胞的增殖作用;(4)Cer也可通过某种机制促进外源PA、lysoPA、SPP、Cer-1-P的降解〔1〕。
, http://www.100md.com
4 Cer信使途径的生物学意义
Cer信使途径是一个普遍存在的、进化上保守的信号传递系统,与cAMP和PI途经相似,大多数哺乳动物细胞可经该途径传递信号〔13〕。有关实验表明该途径参与了细胞生长、增殖、分化、.凋亡、衰老、应激反应及细胞分泌活动等多种生理、病理的过程。总的来看,该途径以传递细胞凋亡等细胞生长抑制活动信号为主,用二酯酰甘油抑制Cer诱导的细胞凋亡,抑制发生在50kbDNA片段期,这种片断是细胞凋亡极早期的图谱特征,表明Cer主要在细胞活动的极早期发挥作用,而细胞外刺激的种类及Cer下游的活动情况将决定其活动的最终结果〔5〕。凋亡作为进化上保守的细胞自杀,在成人组织的早期发育及机体稳态的维持中起重要作用,而许多证据表明Cer是细胞凋亡的关键成分〔10〕。更为重要的是,Cer途径介导的凋亡信号传递受到细胞内抗凋亡体系的严密调控。这种抗凋亡信号体系包括BCL-2体系、DAG-PKC信号途径的活化、p53失活、Cer代谢产物SPP等(比如BCL-2体系中,BCL-2、BCL-XL是抗凋亡成分,而Bax、Bad是促进凋亡的成分)。正是这种抗凋亡和促进凋亡机制之间的平衡状态决定了体内、体外细胞凋亡反应的程度〔13〕。现发现凋亡过程中的许多关键事件集中在线粒体方面:如有实验发现,细胞渗透性Cer诱导的细胞凋亡过程中,在线粒体呼吸链的泛醌位置有ROS产生,并且这种ROS介导了对Cer转录因子NF-κB和AP-1的激活〔14〕;细胞渗透性Cer可刺激GD3神经节苷酯合成,而GD3能够扰乱线粒体膜电位(ΔΨm)而以一种Caspase非依赖性方式诱导细胞凋亡〔15〕;细胞渗透性Cer还可直接抑制线粒体呼吸链的复合体Ⅲ而导致线粒体氧化磷酸化的降低〔16〕。此外,Chmura等人发现,在某些细胞类型中,Cer的产生与否同射线能否诱导细胞凋亡相关:野生型WEHI-231细胞曝露于射线后有nSMase(但不是aSMase)的激活,并发生凋亡,而经过选择的不产生Cer的WEHI-231亚系细胞曝露于射线后没有nSMase的活化和凋亡发生。进一步以外源Cer类似物和SP激酶抑制剂研究证实这种凋亡抗性的细胞对Cer介导的凋亡仍然是敏感的,外源Cer类似物可恢复这细胞的凋亡反应,因此缺乏来自nSMase产生的Cer可能代表了某些肿瘤细胞对治疗产生抗性的机制〔17〕。Michael等也报道Cer是射线诱导凋亡发生的重要中介物,Cer信使途径的缺陷使某些肿瘤细胞对凋亡产生了抗性〔18〕。
, 百拇医药
Cer信使途径也参与了细胞增殖与分化的过程。外源C2-Cer或其它水溶性Cer类似物处理HL-60细胞可模拟IFN-γ、1,25-(OH)2-D3的效应而有效地诱导这类细胞向单核细胞分化〔19〕。Jung等报道人工合成的细胞渗透性Cer类似物显示出浓度依赖性地抑制角化细胞DNA合成而导致细胞增殖的抑制,并且发现由这类角化细胞自身合成的TNF-α可增加内源性Cer水平而抑制细胞增殖〔20〕。
此外,Cer信使途径与疾病的发生、发展密切相关。有人报道Cer通过ERK和SAPK途径而增强基质金属蛋白酶1(MMP-1)基因的表达。MMP-1属于MMPS家族成员之一,主要来源于纤维母细胞,能够降解自身的Ⅰ,Ⅱ,Ⅲ,Ⅴ型胶原蛋白。已知MMPs在血管形成、组织修复、炎症疾病时过多的结缔组织的降解中起重要作用;而更为重要的是,MMPS对组织基底膜和胞外基质的降解在肿瘤细胞的侵袭和转移中起关键作用〔21〕。
, http://www.100md.com 5 小结
对Cer信使途径的研究在过去的几年当中有了很大的进展,许多新的效应分子得到了确认。然而值得注意的是,目前大多数实验结果是经应用人工合成的短链Cer类似物处理细胞并经较长时间作用后得到的,可能未反映细胞对外来刺激而在生理上短暂产生的少量天然Cer所具有的效应,而且合成性Cer类似物加入完整细胞中有可能转变为其它具有生物活性的鞘脂物质,从而经其它信号途径影响细胞功能〔22〕。当前主要集中在对调控Cer生成的酶及Cer效应机制的研究上〔23〕,还有很多问题有待阐明。比如对研究较为深入的Cer介导的凋亡信号传递,虽然确立了几条可能存在的途径,但是否还有其它分子参与,各分子之间相互作用的机制,最终又启动了那些基因的表达,尤其是该途径与其它胞内信使途径之间的相互作用位点,所有这些问题均有待深入研究。
〔参考文献〕
〔1〕 G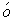 mez-Muňoz A. Modulation of cell signalling by ceramides〔J〕. Bioc-him Biophys Acta,1998,391:109.
mez-Muňoz A. Modulation of cell signalling by ceramides〔J〕. Bioc-him Biophys Acta,1998,391:109.
, http://www.100md.com
〔2〕 Liu B,Hannun YA. Inhibition of the neutral magnesium-dependent sphingomyelinase by glutathione〔J〕. J Biol Chem,1997,272(26):16281~16287.
〔3〕 Tomiuk S,Hofmann K ,Nix M,et al. Cloned mammalian neutral sphingomyelinase:functions in sphingolipid signalling?〔J〕. Proc Natl Acad Sci USA,1998,95:3638~3643.
〔4〕 Garzotto M,White-Jones M,Jiang YW,et al. 12-O-tetradelanoylphorb-ol-13-acetate-induced apoptosis in LN-CaP cells is mediated through ceramide synthase〔J〕. Cancer Res,1998;58(10):2260~2264.
, 百拇医药
〔5〕 丁振华.第二信使神经酰胺〔J〕.生命的化学,1997,17(4):27.
〔6〕 Jarvis WD,Fornari FA,Auer Kl,et al. Coordinate regulation of stress-and mitogen-activated protein kinase in the apoptotic actions of ceramide and sphingosine〔J〕. Mol Pharmacol,1997,52(6):935~947.
〔7〕 Ohmori M,Shirasawu S,Furuse M,et al. Activated Ki-ras enhances sensitivity of ceramide-induced apoptosis without c-Jun NH2-terminal kinase/stess-activated protein or extracellular signal-regulated kinase activation in human colon cancer cells〔J〕. Cancer Res,1997,57(21):4714~4717.
, http://www.100md.com
〔8〕 Shirakabe K,Yamaguchi K,Shibuya H,et al. TAK1 mediates the ceramide signalling to stress-activated protein kinase/c-Jun N-terminal kinase〔J〕. J Biol Chem,1997,272(13):8141~8144.
〔9〕 Shibuya H,Iwata H,Masuyama N,et al. Role of TAK1 and TAB1 in BMP sigmalling in early xenopus development〔J〕. EMBO J,1998,17(4):1019~1028.
〔10〕 Cuvillier O,Rosenthal DS,Smulson ME,et al. Sphigosine 1-phosphate inhibits activation of caspases that cleave poly(ADP-ribose)polymerase and lamins during Fas-and ceramide-me-diated apoptosis in Jurkat T lymphocytes〔J〕. J Biol Chem,1998,273(5):2910~2916.
, 百拇医药
〔11〕 Thornberry NA. The caspase family of cysteine proteases〔J〕. Br Med Bull,1996,53(3):478~490.
〔12〕 Kuo ML,Chen CW,Jee SH,et al. Transforming growth factor beta1 attenuates ceramide-induced CPP32/Yama activation and apoptosis in human leukamia HL-60 cells〔J〕. Biochem J 1997,327(pt3):663~667.
〔13〕 Haimovitz-Friedman A,Kolesnick RN,Fuks Z. Ceramide signaling in apoptosis〔J〕. Br Med Bull,1997,53(3):539~553.
, 百拇医药
〔14〕 Quillet-Mary A,Jaffrezou JP,Mansat V,et al. Implication of mitochondrial hydrogen peroxide generation in ceramide-induced apoptosis〔J〕. J Biol Chem,1997,272(34):21388~21395..
〔15〕 DeMaria-R,Lenti L,Malisan F,et al. Requirement for GD3 ganglioside in CD95-and ceramide-induced apoptosis〔J〕. Science,1997,277(5332):1652~1655.
〔16〕 Gudz TI,Tserng KY,Hoppel CL. Direct inhibition of mitochondrial respiratory chain complex Ⅲ by cell-perme-able ceramide〔J〕. J Biol Chem,1997,272(39):24154~24158.
, 百拇医药
〔17〕 Chmura SJ,Nodzenski E,Beckett MA,et al. Loss of ceramide production confers resistance to radiation-induced apoptosis〔J〕. Cancer Res,1997,57:1270~1275.
〔18〕 Michael JM,Lavin MF,Watters DJ. Resistance to radiation-induced apoptosis in Burkitt's lymphoma cells is associated with defective ceramide signaling〔J〕. Cancer Res,1997,57(16):3600~3605.
〔19〕 Hannun YA. The sphingomyelin cycle and the second messenger function of ceramide〔J〕. J Biol Chem,1994,269(5):3125~3128.
, http://www.100md.com
〔20〕 Jung EM,Griner RD,Mann-Blakeney R,et al. A potential role for ceramide in the regulation of mouse epidermal keratinocyte proliferation and differentiation〔J〕. J Invest Dermatol,1998,110(4):318~323.
〔21〕 Reunanen N,Westermarck J,Hakkinen L,et al. Enhancement of fibroblast collagenase (matrix metalloproteinase-1) gene expression by ceramide is mediated by extracellular signal regulated and stress-activated protein kinase pathways〔J〕. J Biol Chem,1998,273(9):5137~5145.
〔22〕 Kolesnick RN,Kronke M. Regulation of ceramide production and apoptosis〔J〕. Annu Rev Physiol,1998,60:643~665.
〔23〕 HannunYA,Obeid LM. Cermide and the eukaryotic stress response〔J〕. Biochem Soc Trans,1997,25:1171~1175.
收稿日期:1999-01-11;修回日期:1999-02-05, 百拇医药
单位:中山医科大学肿瘤研究所治疗基础室,(广州,510060)
关键词:信号传递;神经酰胺;生物学意义
癌症990539
曾益新(审校)
中图分类号:R-05;R329.28;R344 文献标识码:A
文章编号:1000-467X(1999)05-0605-03
(神经)鞘磷脂(Sphingomyelin,SM)长期以来被认为只是生物膜的一个结构成分,主要分布在脂双层的外层,不知它有什么功能。然而,自从1991年Kolesnick等首先报道甘油二酯(DAG)可诱导SM水解生成神经酰胺(Ceramide,Cer),并发现Cer本身不能象DAG那样激活PKC以来,Cer作为胞内第二信使得到了广泛的证实。作为第二信使,Cer在介导细胞生长、增殖、分化、凋亡及损伤等多种生理、病理过程中起着广泛的作用。本文就Cer生成相关的酶、Cer的效应分子及其与糖脂和其它鞘脂代谢物[如鞘氨醇(Sphingosine)、鞘氨醇1磷酸(Sphingosine-1-Phospha-te)]介导的信号传递途径之间的联系作一综述,并简述Cer信使途径的生物学意义。
, 百拇医药
1 参与Cer生成的酶
1.1 鞘磷脂酶(Sphingomyelinas-e,SMase)
SM可在SMase催化下水解产生Cer和磷酸胆碱,此过程是可逆的,因此称为SM循环。多种激活因素(如TNF-α、IFN-γ、ILs、FasL、1,25-(OH)2-D3、地塞米松、化疗药物、电离辐射及热刺激等)可诱导诸如U937、HL-60、JurkatT细胞、Molt4、PC12等细胞内SM水解而产生Cer,最终导致细胞周期停滞或凋亡等现象的发生。根据SMase的体外活化条件及最适pH值的不同,现发现至少存在4种SMase:(1)膜型中性SMase(nSMase),其活性依赖于Mg2+及中性pH环境(最适pH7.0~7.5),(2)胞浆型nSMase,为Mg2+非依赖性。BinLiu等报道该酶可被生理浓度的GSH抑制,GSH合成抑制剂可导致时间依赖性的GSH降低,而在许多细胞损伤和凋亡过程中发现有GSH的减少,提示GSH的降低可能是nSMase活化的重要机制。(3)酸性SMase(aSMase),主要分布于溶酶体及核内体,激活时需DAG,最适pH值4.5~5.0。(4)碱性SMase,激活时需胆盐,最适pH值9.0。前三种参与信号传递已得到确认,碱性SMase是否与信号传递相关有待进一步研究证实,其中nSMase被认为是SM循环的调节性激活和信号传递中的关键性酶。人的nSMase和aSMase已被克隆,其中nSMase为47.6KD的糖蛋白,由423个氨基酸残基组成。至今测得的具有最强的nSMase活性的组织是中枢神经系统,这与nSMase的广泛组织分布不相一致,提示可能存在一个脑组织特异性的nSMase的活化过程〔1~3〕。
, 百拇医药
1.2 Cer酶(Ceramidase)和Cer合酶(Ceramide Synthase)
Cer在Ceramidase催化下脱酰基生成鞘氨醇,鞘氨醇也可在Cer合酶催化下生成Cer,因此这二种酶在控制细胞内Cer水平起重要作用。Cer可通过激活Cer合酶而从头合成,但至今未发现鞘氨醇头合成途径,因此体内鞘氨醇的生成必需依赖于Ceramidase。许多实验发现Cer和鞘氨醇常诱导相反或不同的生物效应,因此这两种可相互转变的鞘脂代谢物生成的平衡对调控细胞功能可能是一个重要因素。此外,发现诱导细胞凋亡发生的Cer不一定只来自SM的水解:抗癌药物daunorubicin可刺激P388和U937细胞内Cer水平升高并发生凋亡,但此种Cer是通过激活Cer合酶而从头合成的,这是从研究fumonisins得到的结果。Fumonisins是一类Cer合酶的天然的、特异性抑制物,FumonisinB1(FB1)可抑制双氢鞘氨醇-N-酰基转移酶(Sphinganine N-acyltransferase),即Cer合酶的活性而阻断daunorubicin引发的Cer升高及凋亡。Garzotto等报道TPA通过激活PKC-α而诱导LNCaP前列腺细胞凋亡是由Cer合酶介导的,因为此过程中aSMase和nSMase活性均未见升高,并且FB1可取消TPA诱导的Cer产生和细胞凋亡〔1,4〕。
, 百拇医药
1.3 磷脂酸磷酸水解酶(phosphatidate phohydrolase,PAP)
PAP以两种形式存在,PAP-1存在于胞浆,为Mg2+依赖性,可转移至内质网或线粒体,也可转移至细胞膜内表面,并降解由PLD活化产生的磷脂酸(PA)而生成DAG,但PAP-1是否参与信号传递有待证实〔1〕。PAP-2是一膜结合蛋白,为Mg2+非依赖性,也可降解PA产生DAG,但这种PLD来源的DAG不能激活PKC,因此PAP-2可能是通过PA而终止信号传递。PAP-2也可降解Cer-1-P和Sph-1-P(SPP),因此PAP-2可通过降解这类物质而终止来自PA、Cer-1-P、Sph-10-P的信号,但也会产生DAG、Cer和Spingosine(SP)一类具有生物活性的物质,可见PAP可通过调节这类参与细胞活化和控制细胞生长的生物活性脂质之间的平衡而在细胞信号传递中起重要作用〔1〕。
, http://www.100md.com
2 参与Cer信号传递的效应分子
Cer作为第二信使,必定要通过一系列效应分子的级联放大反应,最终激活核内相关基因而产生不同的生物学效应。目前发现神经酰胺活化的蛋白磷酸酶(Ceramide-activated protein phosphatase,CAPP)、神经酰胺活化的蛋白激酶(Ceramide-activated profein kinase,CAPK)、PKC-ζ及原癌基因产物VaV等是Cer的直接靶分子〔5〕。此外,最近在不同细胞系中还发现有下列几种效应分子参与Cer介导的信号传递。
2.1 JNK和ERK
JNK(C-Jun N-terminal Kinase),又称SAPK(应激活化的蛋白激酶),是MAPK超家族的亚族成员,JNK介导的信号传递通路与细胞生长抑制、凋亡及炎症反应相关;ERK(extracellular signal-regulated kinase)途径主要与细胞增殖相关,生长因子和鞘氨醇等可激活ERK而促进细胞生长。实验资料表明,Cer可迅速激活P46-JNK1/P54-JNK2,从而增强JNK的主要底物C-Jun的表达和活性增高,同时Cer可使P42-ERK1/P44-ERK2活性逐渐地、中等程度地降低,而且发现JNK的底物C-Jun的表达和活性的增强对Cer诱导的细胞凋亡发生是绝对必需的〔6〕。但有人在以C2-Cer处理含有Ki-ras突变的人HCT-116结肠癌细胞研究中发现,尽管C2-Cer引起该细胞显著凋亡,但没有观察到JNK/SAPK的激活,可能在该类细胞中有其他信号途径参与了Cer诱导的凋亡〔7〕。
, 百拇医药
2.2 TAK1
TAK1(TGF-βactivated kinase),为丝氨酸/苏氨酸蛋白激酶,属于MAPKKK家族成员,人和鼠的TAK1氨基酸序列高度同源(同源性达98%),因此TAK1是进化上相当保守的激酶。据报道,Cer可直接激活TAK1,活化的TAK1导致SEK1/MKK4激活,后者可继续活化JNK/SAPK,而TAK1的非活化型阻止Cer诱导的JNK/SAPK的激活,说明TAK1是Cer活化JNK/SAPK的关键性中间物,其它脂类物质(包括DAG,花生四烯酸)处理细胞时发现TAK1未被激活,提示TAK1参与了Cer诱发的信号传递〔8〕。此外,还发现TAK1结合蛋白(TAB1)的过表达几乎完全抑制TGF-β刺激TAK1的活化,但对Cer诱导的TAK1激活作用甚微,因此Cer对TAK1的激活是TAB1非依赖性的〔11〕。
2.3 Caspases
, http://www.100md.com
Caspases,属于天冬氨酸特异的半胱氨酸蛋白酶家族,目前根据其种系发生可分为三个亚族:(1)CED-3亚族,包括Caspase-3/CPP32、Caspase-6、Caspase-7、Caspase-8、Caspase-9和Caspase-10;(2)ICE家族,包括Caspase-1、Caspase-4、Caspase-5、Caspase-11;(3)Nedd-2亚族,以Caspase-2为代表。DNA修复酶PARP、DNA-PKcs和核结构蛋白NuMa、lamins等是这类酶的底物,这些底物的水解可能与细胞凋亡时核形态和生化上的改变有关。CED-3亚族的几个Caspase可在凋亡早期催化PARP的分解,其中Caspase-3/CPP32被认为最有效(其次为Caspase-7);核lamins的降解是浓缩染色质进入凋亡小体(凋亡晚期典型的形态结构)所必需的,Caspase-6是目前唯一所知的水解lamins的酶〔10,11〕。Kuo等报道Caspase-3在C2-Cer诱导的HL-60细胞凋亡过程中被激活,而Caspase-3的特异性抑制物Ac-DEVD-CHO可有效地保护Cer诱导的细胞死亡,提示Caspase-3在Cer诱导的凋亡过程中起关键作用〔12〕
, http://www.100md.com
3 Cer信使途径与糖脂及其他鞘脂代谢物介导的信号途径之间的相互作用
许多文献报道了Cer能够诱导细胞凋亡而DAG具有抗凋亡的特性,并且在多种类型细胞中发现因Cer水平增高而诱导的细胞死亡可被DAG所抑制,因此在糖脂和鞘脂信号途经之间必然存在相互作用的位点。目前发现以下事实:(1)细胞渗透性C2和C6-Cer可抑制诸如血清、凝血酶、磷脂酸、SPP或佛波酯等PLD(磷脂酶D)激活剂刺激的PLD活性的增加,而双氢Cer对此无多大影响,说明Cer作用具有特异性;(2)PKC是一类由多种同功酶组成的结构相关的丝氨酸/苏氨酸蛋白酶激酶,与许多细胞功能相关。目前只发现PKC-α、δ、ε受Cer调节。最近有报道称Cer可诱导PKC-δ、ε从胞膜向胞浆的转位,并且外源SMase、TNF-α、Fas抗体(均可诱导凋亡发生)有类似功能,提示PKC-δ、ε的转位在Cer介导的凋亡中起重要作用。(3)细胞渗透性Cer抑制PA、lysoPA、SPP、C2-/C8-Cer-1-P及天然Cer-1-P刺激的DNA合成,即抑制了这类物质对细胞的增殖作用;(4)Cer也可通过某种机制促进外源PA、lysoPA、SPP、Cer-1-P的降解〔1〕。
, http://www.100md.com
4 Cer信使途径的生物学意义
Cer信使途径是一个普遍存在的、进化上保守的信号传递系统,与cAMP和PI途经相似,大多数哺乳动物细胞可经该途径传递信号〔13〕。有关实验表明该途径参与了细胞生长、增殖、分化、.凋亡、衰老、应激反应及细胞分泌活动等多种生理、病理的过程。总的来看,该途径以传递细胞凋亡等细胞生长抑制活动信号为主,用二酯酰甘油抑制Cer诱导的细胞凋亡,抑制发生在50kbDNA片段期,这种片断是细胞凋亡极早期的图谱特征,表明Cer主要在细胞活动的极早期发挥作用,而细胞外刺激的种类及Cer下游的活动情况将决定其活动的最终结果〔5〕。凋亡作为进化上保守的细胞自杀,在成人组织的早期发育及机体稳态的维持中起重要作用,而许多证据表明Cer是细胞凋亡的关键成分〔10〕。更为重要的是,Cer途径介导的凋亡信号传递受到细胞内抗凋亡体系的严密调控。这种抗凋亡信号体系包括BCL-2体系、DAG-PKC信号途径的活化、p53失活、Cer代谢产物SPP等(比如BCL-2体系中,BCL-2、BCL-XL是抗凋亡成分,而Bax、Bad是促进凋亡的成分)。正是这种抗凋亡和促进凋亡机制之间的平衡状态决定了体内、体外细胞凋亡反应的程度〔13〕。现发现凋亡过程中的许多关键事件集中在线粒体方面:如有实验发现,细胞渗透性Cer诱导的细胞凋亡过程中,在线粒体呼吸链的泛醌位置有ROS产生,并且这种ROS介导了对Cer转录因子NF-κB和AP-1的激活〔14〕;细胞渗透性Cer可刺激GD3神经节苷酯合成,而GD3能够扰乱线粒体膜电位(ΔΨm)而以一种Caspase非依赖性方式诱导细胞凋亡〔15〕;细胞渗透性Cer还可直接抑制线粒体呼吸链的复合体Ⅲ而导致线粒体氧化磷酸化的降低〔16〕。此外,Chmura等人发现,在某些细胞类型中,Cer的产生与否同射线能否诱导细胞凋亡相关:野生型WEHI-231细胞曝露于射线后有nSMase(但不是aSMase)的激活,并发生凋亡,而经过选择的不产生Cer的WEHI-231亚系细胞曝露于射线后没有nSMase的活化和凋亡发生。进一步以外源Cer类似物和SP激酶抑制剂研究证实这种凋亡抗性的细胞对Cer介导的凋亡仍然是敏感的,外源Cer类似物可恢复这细胞的凋亡反应,因此缺乏来自nSMase产生的Cer可能代表了某些肿瘤细胞对治疗产生抗性的机制〔17〕。Michael等也报道Cer是射线诱导凋亡发生的重要中介物,Cer信使途径的缺陷使某些肿瘤细胞对凋亡产生了抗性〔18〕。
, 百拇医药
Cer信使途径也参与了细胞增殖与分化的过程。外源C2-Cer或其它水溶性Cer类似物处理HL-60细胞可模拟IFN-γ、1,25-(OH)2-D3的效应而有效地诱导这类细胞向单核细胞分化〔19〕。Jung等报道人工合成的细胞渗透性Cer类似物显示出浓度依赖性地抑制角化细胞DNA合成而导致细胞增殖的抑制,并且发现由这类角化细胞自身合成的TNF-α可增加内源性Cer水平而抑制细胞增殖〔20〕。
此外,Cer信使途径与疾病的发生、发展密切相关。有人报道Cer通过ERK和SAPK途径而增强基质金属蛋白酶1(MMP-1)基因的表达。MMP-1属于MMPS家族成员之一,主要来源于纤维母细胞,能够降解自身的Ⅰ,Ⅱ,Ⅲ,Ⅴ型胶原蛋白。已知MMPs在血管形成、组织修复、炎症疾病时过多的结缔组织的降解中起重要作用;而更为重要的是,MMPS对组织基底膜和胞外基质的降解在肿瘤细胞的侵袭和转移中起关键作用〔21〕。
, http://www.100md.com 5 小结
对Cer信使途径的研究在过去的几年当中有了很大的进展,许多新的效应分子得到了确认。然而值得注意的是,目前大多数实验结果是经应用人工合成的短链Cer类似物处理细胞并经较长时间作用后得到的,可能未反映细胞对外来刺激而在生理上短暂产生的少量天然Cer所具有的效应,而且合成性Cer类似物加入完整细胞中有可能转变为其它具有生物活性的鞘脂物质,从而经其它信号途径影响细胞功能〔22〕。当前主要集中在对调控Cer生成的酶及Cer效应机制的研究上〔23〕,还有很多问题有待阐明。比如对研究较为深入的Cer介导的凋亡信号传递,虽然确立了几条可能存在的途径,但是否还有其它分子参与,各分子之间相互作用的机制,最终又启动了那些基因的表达,尤其是该途径与其它胞内信使途径之间的相互作用位点,所有这些问题均有待深入研究。
〔参考文献〕
〔1〕 G
mez-Muňoz A. Modulation of cell signalling by ceramides〔J〕. Bioc-him Biophys Acta,1998,391:109., http://www.100md.com
〔2〕 Liu B,Hannun YA. Inhibition of the neutral magnesium-dependent sphingomyelinase by glutathione〔J〕. J Biol Chem,1997,272(26):16281~16287.
〔3〕 Tomiuk S,Hofmann K ,Nix M,et al. Cloned mammalian neutral sphingomyelinase:functions in sphingolipid signalling?〔J〕. Proc Natl Acad Sci USA,1998,95:3638~3643.
〔4〕 Garzotto M,White-Jones M,Jiang YW,et al. 12-O-tetradelanoylphorb-ol-13-acetate-induced apoptosis in LN-CaP cells is mediated through ceramide synthase〔J〕. Cancer Res,1998;58(10):2260~2264.
, 百拇医药
〔5〕 丁振华.第二信使神经酰胺〔J〕.生命的化学,1997,17(4):27.
〔6〕 Jarvis WD,Fornari FA,Auer Kl,et al. Coordinate regulation of stress-and mitogen-activated protein kinase in the apoptotic actions of ceramide and sphingosine〔J〕. Mol Pharmacol,1997,52(6):935~947.
〔7〕 Ohmori M,Shirasawu S,Furuse M,et al. Activated Ki-ras enhances sensitivity of ceramide-induced apoptosis without c-Jun NH2-terminal kinase/stess-activated protein or extracellular signal-regulated kinase activation in human colon cancer cells〔J〕. Cancer Res,1997,57(21):4714~4717.
, http://www.100md.com
〔8〕 Shirakabe K,Yamaguchi K,Shibuya H,et al. TAK1 mediates the ceramide signalling to stress-activated protein kinase/c-Jun N-terminal kinase〔J〕. J Biol Chem,1997,272(13):8141~8144.
〔9〕 Shibuya H,Iwata H,Masuyama N,et al. Role of TAK1 and TAB1 in BMP sigmalling in early xenopus development〔J〕. EMBO J,1998,17(4):1019~1028.
〔10〕 Cuvillier O,Rosenthal DS,Smulson ME,et al. Sphigosine 1-phosphate inhibits activation of caspases that cleave poly(ADP-ribose)polymerase and lamins during Fas-and ceramide-me-diated apoptosis in Jurkat T lymphocytes〔J〕. J Biol Chem,1998,273(5):2910~2916.
, 百拇医药
〔11〕 Thornberry NA. The caspase family of cysteine proteases〔J〕. Br Med Bull,1996,53(3):478~490.
〔12〕 Kuo ML,Chen CW,Jee SH,et al. Transforming growth factor beta1 attenuates ceramide-induced CPP32/Yama activation and apoptosis in human leukamia HL-60 cells〔J〕. Biochem J 1997,327(pt3):663~667.
〔13〕 Haimovitz-Friedman A,Kolesnick RN,Fuks Z. Ceramide signaling in apoptosis〔J〕. Br Med Bull,1997,53(3):539~553.
, 百拇医药
〔14〕 Quillet-Mary A,Jaffrezou JP,Mansat V,et al. Implication of mitochondrial hydrogen peroxide generation in ceramide-induced apoptosis〔J〕. J Biol Chem,1997,272(34):21388~21395..
〔15〕 DeMaria-R,Lenti L,Malisan F,et al. Requirement for GD3 ganglioside in CD95-and ceramide-induced apoptosis〔J〕. Science,1997,277(5332):1652~1655.
〔16〕 Gudz TI,Tserng KY,Hoppel CL. Direct inhibition of mitochondrial respiratory chain complex Ⅲ by cell-perme-able ceramide〔J〕. J Biol Chem,1997,272(39):24154~24158.
, 百拇医药
〔17〕 Chmura SJ,Nodzenski E,Beckett MA,et al. Loss of ceramide production confers resistance to radiation-induced apoptosis〔J〕. Cancer Res,1997,57:1270~1275.
〔18〕 Michael JM,Lavin MF,Watters DJ. Resistance to radiation-induced apoptosis in Burkitt's lymphoma cells is associated with defective ceramide signaling〔J〕. Cancer Res,1997,57(16):3600~3605.
〔19〕 Hannun YA. The sphingomyelin cycle and the second messenger function of ceramide〔J〕. J Biol Chem,1994,269(5):3125~3128.
, http://www.100md.com
〔20〕 Jung EM,Griner RD,Mann-Blakeney R,et al. A potential role for ceramide in the regulation of mouse epidermal keratinocyte proliferation and differentiation〔J〕. J Invest Dermatol,1998,110(4):318~323.
〔21〕 Reunanen N,Westermarck J,Hakkinen L,et al. Enhancement of fibroblast collagenase (matrix metalloproteinase-1) gene expression by ceramide is mediated by extracellular signal regulated and stress-activated protein kinase pathways〔J〕. J Biol Chem,1998,273(9):5137~5145.
〔22〕 Kolesnick RN,Kronke M. Regulation of ceramide production and apoptosis〔J〕. Annu Rev Physiol,1998,60:643~665.
〔23〕 HannunYA,Obeid LM. Cermide and the eukaryotic stress response〔J〕. Biochem Soc Trans,1997,25:1171~1175.
收稿日期:1999-01-11;修回日期:1999-02-05, 百拇医药