大鼠外侧丘系核GABA能免疫阳性神经元向下丘的投射①
作者:李佩英 岳毅刚 骆耐香 陆明深 张德兴
单位:李佩英 骆耐香 陆明深 桂林医学院解剖教研室 广西桂林市 541001;岳毅刚 桂林医学院整形外科 张德兴 广东药学院基础部
关键词:听觉中枢;下丘;外侧丘系核;GABA能投射;大鼠
华夏医学990601 摘要 用HRP逆行追踪结合抗GABA的免疫细胞化学双标记法对外侧丘系核 (NLL)向下丘中央核(ICC)的GABA能投射进行了研究。结果:在双侧外侧丘系背侧核(DNLL)及同侧外侧丘系中间核(INLL)和外侧丘系腹侧核(VNLL)均可见到大量HRP单标,GABA单标及HRP/GABA双标细胞。其中HRP/GABA双标细胞在同侧VNLL最多(41%),同侧INLL为21%,同侧DNLL为8%,对侧DNLL为29%,各亚核双标细胞占投射神经元的比例:同侧VNLL为70%,同侧INLL为63%,同侧DNLL为51%,而对侧DNLL为75%。对侧INLL及VNLL未见HRP单标及HRP/GABA双标细胞。以上结果表明,双侧DNLL、同侧INLL和VNLL存在着大量向ICC投射GABA能神经元。本研究提示NLL通过GABA能投射向IC施加抑制性影响。
, 百拇医药
中图分类号 Q421;Q422
The Projection of GABA-immunoreactive Neurons in the Nuclei of
Lateral Lemniscus to the Inferior Colliculus of Rat
Li Peiying,Yue Yigang,Luo Naixiang,et al.
Dept.of Anatomy of Guilin Medical College1 541001
Abstract The GABAergic projection from the nuclei of lateral lemiscus to the inferior colliculus of rat was studied by combining the retrograde transport of horseradish peroxidase(HRP) and immunohistochemistry for GABA.Numerous HRP-labeled cells, GABAergic neurons and HRP/GABA neurons were found in dorsal nucleus of the lateral lemniscus(DNLL)bilaterally, in intermediate nucleus of the lateral lemniscus(INLL) and ventral nucleus of the lateral lemnisus(VNLL)ipsilaterally.The number of double-labeled neurons in ipsilateral DNLL was higher than the other nuclei,reaching approximately 41% of the total number of the double-labeled neurons.Additionally,in the contralateral DNLL,ipsilateral INLL and VNLL,the number was about 21%,8% and 29% respectively.The sum of doubled -labeled neurons of ipsilateral VNLL,INLL,DNLL and contralateral DNLL reached approximately 70%,63%,51% and 75% respectively of their HRP-labeled cells.Any double-labeled neurons in contralateral INLL and VNLL were found.The results revealed that many GABAergic neurons in bilateral DNLL,ipsilateral INLL and VNLL projected to the central nucleus of the inferior colliculus.
, 百拇医药
Key words central audinary system;inferior colliculus;nuclei of the lateral lemniscus;GABAergic projection;rat
下丘(IC)是中脑顶盖的一部分,是重要的听觉中枢,在听觉传导路中处于一个关键的位置,在处理单耳和双耳信息中起着至关重要的作用。可分为三个亚核,即下丘中央核(ICC),下丘外周核(ICX)和下丘背侧核(ICD)。外侧丘系核(NLL)是位于听觉上行通路二级神经元的蜗神经核发出的二级纤维(即外侧丘系)中的几群细胞,沿中脑的外侧缘排列,呈背腹走向,通常可分为外侧丘系背侧核(DNLL),外侧丘系中间核(INLL)及外侧丘系腹侧核(VNLL)三群。
免疫细胞化学研究表明,下丘存在着非常丰富的GABA阳性终末和神经元[1,2]。对这些终末的来源,有人认为是来自于本核内的局部神经元,其轴突仅终止于IC内部的细胞[3],但Winer[2]则推测部分是来自于其它部位向IC的GABA能投射。NLL的三个亚核中许多神经元为GABA和GAD阳性神经元[2]。笔者应用荧光金逆行追踪的方法及一些其它实验业已证明,大鼠NLL发出大量的纤维投射到ICC[4]。那么NLL的GABA能神经元是否投射到ICC?已有实验获得了一些间接证据[5,6],但能提供直接形态学依据的报道还很少见。本实验利用HRP逆行追踪技术结合抗GABA的免疫细胞化学双标记法进行研究,以期为证实NLL-IC的GABA能投射的存在提供直接证据。
, http://www.100md.com
1 材料与方法
实验选用健康成年Wistar大鼠15只,体重200~250g,雌雄不拘。0.4% 戊巴比妥钠腹腔注射麻醉后,将动物固定于立体定位仪上,参照Watson的《大鼠脑立体定位图谱》,用微量注射器将30%HRP水溶解(Sigma Ⅵ)加压注射到IC,注射量为0.3μl。动物存活48h后,再次以戊巴比妥钠麻醉,经升主动脉先用100ml生理盐水快速冲洗血管床,再以预冷的含2%多聚甲醛和0.5%戊二醛的PB液(0.1mol/L,pH7.4)500ml 灌注,留针于心脏1h后灌以预冷的含10%蔗糖的PB液(0.1mol/L,pH7.4)500ml。灌毕取脑,入20%的蔗糖PB液中过夜。次日用振动切片机切片,片厚50μm,隔3片取2片,分二套收集于0.1ml/L的PB液中(pH7.4)。
第一套片:先按TMB法作HRP反应:切片经蒸馏水洗6次后,再经温育液(22℃)预浸20min。温育液的配制:A液由亚硝酰铁氰化钠50mg,醋酸缓冲液(pH3.3)2.5ml,蒸馏水46ml;B液由TMB2.5mg,无水酒精1.25ml加温溶解而成。在临用前将B液加入到A液中,即成温育液。在50ml温育液中, 加入0.3% H2O2 1.25ml,摇匀后放入切片孵育20min,经洗保存液(醋酸缓冲液pH3.3)洗6次,再按Rye法[7]稳定TMB产物,光镜下证实有HRP标记细胞后,再按ABC法作GABA免疫细胞化学染色。免疫细胞化学染色程序:20%正常羊血清孵育切片,室温,30min;兔抗GABA血清(Sigma,1∶100),4℃,72h;羊抗兔IgG(Vector ,1∶200),室温,1h;ABC复合物(Vector, 1∶200),室温,1h;含0.02%DAB和0.003%H2O2的TB缓冲液中呈色,室温,10~30min。除第一步与第三步外,其余各步间均以PBS充分漂洗。切片反应完毕后用PBS洗3次,10min/次。经常规脱水、透明后,用中性树胶封片,光镜观察。
, http://www.100md.com
第二套切片:用做对照试验:①置换试验:用正常羊血清代替抗GABA血清,其余步骤同第一套切片。结果为阴性,说明实验使用的免抗GABA抗体具有特异性。②吸收试验:Sigma公司在鉴定本产品时已做。
2 结果
在15例动物中,选择注射部位准确位于下丘中央核的10侧进行观察。在双侧DNLL和同侧INLL、VNLL中均可见到HRP单标、GABA单标及HRP/GABA双标三种细胞,每例动物选择部位相当的3张切片,共计30张切片进行计数分析,计算每个核团每张切片中平均细胞数。结果见表1。
表1 每张切片HRP、HRP/GABA双标细胞在NLL各亚核中的平均数 (n,%) 核团
HRP
HRP%
, 百拇医药 双标细胞
双标%
双标/HRP%
双侧DNLL
170
17
128
29
75
同侧DNLL
65
10
33
, http://www.100md.com
8
51
同侧INLL
140
22
89
21
63
同侧VNLL
262
41
184
42
, http://www.100md.com 70
总计
637
100
434
100
2.1 HRP标记细胞
在TMB法反应切片上,HRP反应产物呈蓝黑色颗粒,分布于胞浆和树突基部,颗粒之间的胞浆未着色。HRP标记细胞轮廓清晰可见。
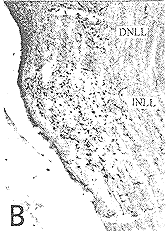
DNLL:在双侧DNLL中均可见于HRP单标细胞(图1、2)。HRP单标细胞大小不等,形态各异,以中等圆形、卵圆形和梭形细胞为主,此外还有少量体积较大的多极细胞,突起清晰可见。梭形细胞的长轴及突起的方向与外侧丘系纤维不平行。对侧DNLL中标记细胞形成同心圆状排列(图1),而同侧DNLL中细胞未排列形成明显的同心圆状(图2)。对侧DNLL中的HRP数量较多,占三个亚核中HRP细胞总数 的27%,而同侧DNLL中HRP数量较少,只占HRP细胞总数的10%。
, http://www.100md.com
图1(A) 对侧DNLL中的HRP单标细胞 ×40
图2(B) 同侧DNLL及INLL的HRP单标细胞 ×40
INLL和VNLL:仅在同侧的INLL和VNLL中观察到HRP细胞(图2,3),体积较DNLL的为小,以中小圆形、卵圆形或长梭形为主,突起较DNLL的为少,细胞长轴多与外侧丘系纤维平行。INLL的HRP细胞较多,占HRP细胞总数的22%(图2),而VNLL的HRP细胞排列密集,数量最多,约占HRP细胞总数的41%(图3)。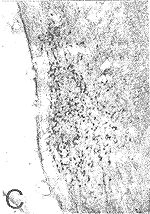
图3(C) 同侧VNLL的HRP单标细胞 ×40
2.2 GABA标记细胞
在低倍镜下,GABA阳性神经元的胞浆呈均匀的棕黄色,在高倍镜下,则可在胞浆中观察到棕黄色的细小颗粒。NLL的三个亚核都有染色强度及数量不等的GABA免疫阳性细胞。其中DNLL中的GABA细胞免疫染色最强,细胞大小不等,形态为多极、梭形、圆形和卵圆形。INLL和VNLL中的 GABA细胞染色较弱,体积较小,为中、小圆形或卵圆形。由于本实验研究的重点在NLL-IC投射细胞,故未对NLL各亚核中GABA细胞进行计数。
, 百拇医药
2.3 HRP/GABA双标细胞
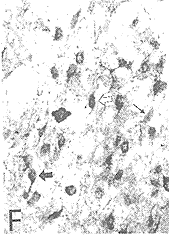
双标记细胞的特点是在均匀的棕黄色背景沉积着蓝黑色的颗粒。在DNLL中(图4),虽然HRP单标细胞形态各异,大小不等,HRP/GABA双标记细胞却以中、小圆形或卵圆形为主,大多极细胞和长梭形细胞却极少被双染。双标细胞数占投射细胞总数的比例,对侧DNLL为75%,同侧DNLL为51%。在INLL和VNLL中也有大量的HRP/GABA双标细胞,以中等圆形或卵圆形为主(图5,6)。双标细胞占投射细胞的比例,INLL为63%,而VNLL为70%。各亚核的双标细胞数占三个亚核总双标细胞数的比例,对侧DNLL为 29%,同侧DNLL为8%,同侧INLL为21%,同侧VNLL为42%。

, http://www.100md.com
图4(D) 对侧DNLL中的HRP单标细胞( )、GABA单标细胞(↑)和HRP/GABA双标细胞(
)、GABA单标细胞(↑)和HRP/GABA双标细胞( ) ×160
) ×160
图5(E) 同侧INLL中的HRP单标细胞()、GABA单标细胞(↑)和HRP/GABA双标细胞() ×200
图6(F) 同侧VNLL中的HRP单标细胞()、GABA单标细胞(↑)和HRP/GABA双标细胞() ×200
, http://www.100md.com
3 讨论
3.1 DNLL-IC的GABA能投射
既往的实验表明,DNLL发出纤维支配双侧IC,对侧支配强于同侧[4]。本实验将HRP注入到一侧ICC后,在双侧DNLL观察到大量HRP单标细胞,对侧DNLL内的单标细胞约为同侧的2.7倍,结果与前人的研究一致。有人发现DNLL-IC投射的终末内具有扁平和多形两种囊泡,而这两种囊泡被普遍认为是抑制性突触的特征性结构[5]。Shneiderman[6]将海人酸注入到一侧DNLL后,观察到双侧 IC的GABA释放减少,以对侧为显著。免疫细胞化学实验也显示,DNLL中大部分神经元均为GABA或GAD阳性神经元[1,2]。以上研究均提示DNLL内可能存在着向IC投射的GABA能神经元。本实验用HRP逆行追踪与免疫细胞化学双标记法研究,发现双侧DNLL的许多神经元为HRP/GABA双标神经元,对侧约为同侧的3.6倍,因而证实了DNLL-IC投射神经元中有许多是GABA能神经元,即DNLL中许多神经元是以GABA作为神经递质对IC产生抑制性影响,而且对侧强于同侧,从而为以上实验的推测提供了直接的形态学依据。另外,双标细胞主要为中、小圆形细胞,大多极细胞和长梭形细胞很少被双染,这与De Xingzhang[8]的研究结果一致,提示在DNLL内部,不同形态的神经元可能执行不同的功能。
, http://www.100md.com
关于DNLL-IC的GABA能神经元的机能意义,有些生理学实验进行了研究。用利多卡因阻断NLL-ICC传导后,双侧IC的双音抑制现象有所减弱[9];向DNLL内注入兴奋性氨基酸海人酸或电刺激DNLL可增加对侧ICC的声音诱发抑制反应[9];向一侧DNLL注入兴奋性氨基酸拮抗剂犬尿喹啉酸,对侧ICC的双音抑制作用大为减弱, 而且对两耳声音的时间差及强度差的感受能力也将改变[10]。以上证据表明IC的双音抑制部分由对侧DNLL-IC的 GABA能投射所调节,这种调节可能是通过形成前馈性抑制通路来实现[11]。下丘内这些对双音刺激起反应的神经元,对两耳声音的时间差和强度差反应敏感,而声源定位能力则取决于两耳声音的时间差及强度差 。Kelly[12]用海人酸毁损一侧或两侧DNLL或手术横断Probst连合纤维(为由DNLL投射到对侧DNLL和IC的纤维),可导致声源定位缺失及最小听角上抬,提示交叉的NLL-IC的GABA能投射在提高ICC对双音刺激的敏感性及提高声源定位能力方面起着重要作用。
, 百拇医药
本实验的数据显示在对侧DNLL和同侧DNLL中分别约有75%和50%的投射神经元为双标神经元,这意味着还有少量其它性质的神经元。根据Winer[2]的研究结果,DNLL中没有甘氨酸能神经元,因而DNLL-IC投射为甘氨酸能的可能性不大。而甘氨酸和GABA为中枢神经系统中两种最主要的抑制性递质,因此推测这些非GABA能的投射可能为兴奋性的。Shneiderman[5]发现同侧DNLL-IC投射神经元的轴突终未中有圆形囊泡,这种形态的囊泡被普遍认为含有兴奋性递质;DNLL的神经元细胞内记录显示当电刺激Probst纤维时,可引发兴奋性突触后电位[13]。以上实验均支持DNLL-IC投射中除了大部分是以GABA为递质的抑制性投射以外,少数为兴奋性投射的推测。至于这些兴奋性投射是以哪种物质作为递质,还有待于深入研究。
3.2 INLL-IC和VNLL-IC的GABA能投射
本实验在INLL和VNLL二核仅于同侧观察到大量的HRP单标细胞,其中VNLL的HRP细胞数量为INLL的1.9倍,为同侧DNLL的4倍,为对侧DNLL的1.5 倍。表明此二核发出大量纤维投射至同侧IC,而同侧VNLL是NLL-IC投射的最大来源,符合前人的研究结果。在同侧IVLL和VNLL中还观察到大量的HRP/GABA双标细胞,表明这些核团投射到IC的神经元中有许多为GABA能神经元,其中INLL的双标细胞数占NLL核双标细胞总数的21%,而VNLL为42%,表明同侧VNLL是NLL-IC的GABA能投射的最大来源。INLL中的双标细胞数占HRP细胞数的63%,VNLL则为70%,表明INLL-IC及VNLL-IC投射神经元中也可能有其它性质的神经元存在。Winer[2]的研究结果显示,INLL和VNLL中甘氨酸能神经元的数量比GABA能神经元的多,Saint-Marie[14]根据[3H]标记的甘氨酸作放射自显影研究结果,推测VNLL中GABA能神经元与甘氨酸能神经元共存。故尽管本实验结果显示VNLL-IC和INLL-IC投射神经元中大部分为GABA能,但不排除存在着甘氨酸能的VNLL-IC和INLL-IC投射,也不排除这两种递质共存于同一投射神经元的可能。最后的定论有待于进一步的实验研究,其机能意义也有待进一步探讨。
, 百拇医药
① 基金项目:广西青年科学基金资助课题(No.9631009)。
作者简介:李佩英(1962-),女,1984年毕业于广西医科大学系,获学士学位,1997年在广西医科大学获神经解剖学硕士学位。现任桂林医科学院解剖教研室主任、副教授。
参考文献
1 李佩英,岳毅刚,张德兴,等.树 听觉中枢GABA能神经元及终末的分布. 桂林医学院学报,1996,9(3):229~232
听觉中枢GABA能神经元及终末的分布. 桂林医学院学报,1996,9(3):229~232
2 Winer JA,Larue DT,Pollak GD.GABA and glycine in the central auditory system of the mustache bat:structural substrates for inbibitory neuronal organization.J Comp Nenrol,1995,355:317~353
, http://www.100md.com
3 Douglas L Oliver,Jeffery AW,Gretchen E Beckius,et al. Morphology of GABAergic neurons in the inferior colliculus of the cat.J Comp Neurol,1994,340:27~42
4 Merchan MA,Saldana E,Plaza I.Dorsal nucleus of the lateral lemniscus in the rat :concentric organization and tonotopic projection to the inferior colliculus.J Comp Neurol, 1994,342:259~278
5 Shneiderman A,Oliver DL.EM autoradiographic study of the projections from the dorssal nucleus of the lateral lemniscus:a possible source of inhibitory inputs to the inferior colliculus.J Comp Neuro,1989,286:28~47
, 百拇医药
6 Shneiderman A,Chase MB,Rockwood JM,et al.Evidence for a GABAergic projection from the dorsal nucleus of the lateral lemniscus to the inferior colliculus.J Neurochem, 1993,60:72~82
7 Rye.Stabilization of the tetramethylbenzidine (TMB) reaction product:Application for retrograde and anterograde tracing and combination with immunohistochemistry.J Histochem Cytochem,1984,32(11):1145~1150
8 De Xingzhang,Liang Li,Jack B Kell,et al.GABAergic projections from the lateral lemniscus to the inferior colliculus of the rat.Hear Res,1998,117:1~12
, 百拇医药
9 Faingold CL,Boersma-Anderson CA,Randal ME.Stimulation or blockade of the dorsal nucleus of the lateral lemniscus alters binaural and tonic inhibition in contralateral inferior colliculus neurons.Hear Res,1993,69:98~106
10 Kelly JB,Li L.Two soures of inhibition affecting binaural evoked responses in the rat's inferior colliculus:the dorsal nucleus of the lateral lemniscus and the superior olivary complex.HearRes,1997,104:112~126
, http://www.100md.com 11 Shneiderman A,Oliver DL,Henkel C.Connections of the dorsal nucleus of the lateral lemniscus:an inhitibory parallel pathway in the ascending auditory system? J Comp Neurol,1988,278:188~208
12 Kelly JB,Li L,Van Adel B.Sound Localization after kainic acid lesions of the dorsal nucleus of the lateral lemniscus in the albino rat.Behav Neurosci,1996,110: 1445~1455
13 Wu SH,Kelly JB.In vitro brain slicestudies of the rat's dorsal nucleus of the lateral lemniscus I. Membrane and synaptic response properties.J Neurophysiol,1995,73:780~793
14 Saint Marie RL,Baker RA.Neurotransmitter-specific uptake and retrograde transport of [3H] glycine from the inferior colliculus by ipsilateral projections of the superior olivary complex and nuclei of the lateral lemniscus.Brain Res,1990, 524:244~253
收稿 1999-12-01, 百拇医药
单位:李佩英 骆耐香 陆明深 桂林医学院解剖教研室 广西桂林市 541001;岳毅刚 桂林医学院整形外科 张德兴 广东药学院基础部
关键词:听觉中枢;下丘;外侧丘系核;GABA能投射;大鼠
华夏医学990601 摘要 用HRP逆行追踪结合抗GABA的免疫细胞化学双标记法对外侧丘系核 (NLL)向下丘中央核(ICC)的GABA能投射进行了研究。结果:在双侧外侧丘系背侧核(DNLL)及同侧外侧丘系中间核(INLL)和外侧丘系腹侧核(VNLL)均可见到大量HRP单标,GABA单标及HRP/GABA双标细胞。其中HRP/GABA双标细胞在同侧VNLL最多(41%),同侧INLL为21%,同侧DNLL为8%,对侧DNLL为29%,各亚核双标细胞占投射神经元的比例:同侧VNLL为70%,同侧INLL为63%,同侧DNLL为51%,而对侧DNLL为75%。对侧INLL及VNLL未见HRP单标及HRP/GABA双标细胞。以上结果表明,双侧DNLL、同侧INLL和VNLL存在着大量向ICC投射GABA能神经元。本研究提示NLL通过GABA能投射向IC施加抑制性影响。
, 百拇医药
中图分类号 Q421;Q422
The Projection of GABA-immunoreactive Neurons in the Nuclei of
Lateral Lemniscus to the Inferior Colliculus of Rat
Li Peiying,Yue Yigang,Luo Naixiang,et al.
Dept.of Anatomy of Guilin Medical College1 541001
Abstract The GABAergic projection from the nuclei of lateral lemiscus to the inferior colliculus of rat was studied by combining the retrograde transport of horseradish peroxidase(HRP) and immunohistochemistry for GABA.Numerous HRP-labeled cells, GABAergic neurons and HRP/GABA neurons were found in dorsal nucleus of the lateral lemniscus(DNLL)bilaterally, in intermediate nucleus of the lateral lemniscus(INLL) and ventral nucleus of the lateral lemnisus(VNLL)ipsilaterally.The number of double-labeled neurons in ipsilateral DNLL was higher than the other nuclei,reaching approximately 41% of the total number of the double-labeled neurons.Additionally,in the contralateral DNLL,ipsilateral INLL and VNLL,the number was about 21%,8% and 29% respectively.The sum of doubled -labeled neurons of ipsilateral VNLL,INLL,DNLL and contralateral DNLL reached approximately 70%,63%,51% and 75% respectively of their HRP-labeled cells.Any double-labeled neurons in contralateral INLL and VNLL were found.The results revealed that many GABAergic neurons in bilateral DNLL,ipsilateral INLL and VNLL projected to the central nucleus of the inferior colliculus.
, 百拇医药
Key words central audinary system;inferior colliculus;nuclei of the lateral lemniscus;GABAergic projection;rat
下丘(IC)是中脑顶盖的一部分,是重要的听觉中枢,在听觉传导路中处于一个关键的位置,在处理单耳和双耳信息中起着至关重要的作用。可分为三个亚核,即下丘中央核(ICC),下丘外周核(ICX)和下丘背侧核(ICD)。外侧丘系核(NLL)是位于听觉上行通路二级神经元的蜗神经核发出的二级纤维(即外侧丘系)中的几群细胞,沿中脑的外侧缘排列,呈背腹走向,通常可分为外侧丘系背侧核(DNLL),外侧丘系中间核(INLL)及外侧丘系腹侧核(VNLL)三群。
免疫细胞化学研究表明,下丘存在着非常丰富的GABA阳性终末和神经元[1,2]。对这些终末的来源,有人认为是来自于本核内的局部神经元,其轴突仅终止于IC内部的细胞[3],但Winer[2]则推测部分是来自于其它部位向IC的GABA能投射。NLL的三个亚核中许多神经元为GABA和GAD阳性神经元[2]。笔者应用荧光金逆行追踪的方法及一些其它实验业已证明,大鼠NLL发出大量的纤维投射到ICC[4]。那么NLL的GABA能神经元是否投射到ICC?已有实验获得了一些间接证据[5,6],但能提供直接形态学依据的报道还很少见。本实验利用HRP逆行追踪技术结合抗GABA的免疫细胞化学双标记法进行研究,以期为证实NLL-IC的GABA能投射的存在提供直接证据。
, http://www.100md.com
1 材料与方法
实验选用健康成年Wistar大鼠15只,体重200~250g,雌雄不拘。0.4% 戊巴比妥钠腹腔注射麻醉后,将动物固定于立体定位仪上,参照Watson的《大鼠脑立体定位图谱》,用微量注射器将30%HRP水溶解(Sigma Ⅵ)加压注射到IC,注射量为0.3μl。动物存活48h后,再次以戊巴比妥钠麻醉,经升主动脉先用100ml生理盐水快速冲洗血管床,再以预冷的含2%多聚甲醛和0.5%戊二醛的PB液(0.1mol/L,pH7.4)500ml 灌注,留针于心脏1h后灌以预冷的含10%蔗糖的PB液(0.1mol/L,pH7.4)500ml。灌毕取脑,入20%的蔗糖PB液中过夜。次日用振动切片机切片,片厚50μm,隔3片取2片,分二套收集于0.1ml/L的PB液中(pH7.4)。
第一套片:先按TMB法作HRP反应:切片经蒸馏水洗6次后,再经温育液(22℃)预浸20min。温育液的配制:A液由亚硝酰铁氰化钠50mg,醋酸缓冲液(pH3.3)2.5ml,蒸馏水46ml;B液由TMB2.5mg,无水酒精1.25ml加温溶解而成。在临用前将B液加入到A液中,即成温育液。在50ml温育液中, 加入0.3% H2O2 1.25ml,摇匀后放入切片孵育20min,经洗保存液(醋酸缓冲液pH3.3)洗6次,再按Rye法[7]稳定TMB产物,光镜下证实有HRP标记细胞后,再按ABC法作GABA免疫细胞化学染色。免疫细胞化学染色程序:20%正常羊血清孵育切片,室温,30min;兔抗GABA血清(Sigma,1∶100),4℃,72h;羊抗兔IgG(Vector ,1∶200),室温,1h;ABC复合物(Vector, 1∶200),室温,1h;含0.02%DAB和0.003%H2O2的TB缓冲液中呈色,室温,10~30min。除第一步与第三步外,其余各步间均以PBS充分漂洗。切片反应完毕后用PBS洗3次,10min/次。经常规脱水、透明后,用中性树胶封片,光镜观察。
, http://www.100md.com
第二套切片:用做对照试验:①置换试验:用正常羊血清代替抗GABA血清,其余步骤同第一套切片。结果为阴性,说明实验使用的免抗GABA抗体具有特异性。②吸收试验:Sigma公司在鉴定本产品时已做。
2 结果
在15例动物中,选择注射部位准确位于下丘中央核的10侧进行观察。在双侧DNLL和同侧INLL、VNLL中均可见到HRP单标、GABA单标及HRP/GABA双标三种细胞,每例动物选择部位相当的3张切片,共计30张切片进行计数分析,计算每个核团每张切片中平均细胞数。结果见表1。
表1 每张切片HRP、HRP/GABA双标细胞在NLL各亚核中的平均数 (n,%) 核团
HRP
HRP%
, 百拇医药 双标细胞
双标%
双标/HRP%
双侧DNLL
170
17
128
29
75
同侧DNLL
65
10
33
, http://www.100md.com
8
51
同侧INLL
140
22
89
21
63
同侧VNLL
262
41
184
42
, http://www.100md.com 70
总计
637
100
434
100
2.1 HRP标记细胞
在TMB法反应切片上,HRP反应产物呈蓝黑色颗粒,分布于胞浆和树突基部,颗粒之间的胞浆未着色。HRP标记细胞轮廓清晰可见。
DNLL:在双侧DNLL中均可见于HRP单标细胞(图1、2)。HRP单标细胞大小不等,形态各异,以中等圆形、卵圆形和梭形细胞为主,此外还有少量体积较大的多极细胞,突起清晰可见。梭形细胞的长轴及突起的方向与外侧丘系纤维不平行。对侧DNLL中标记细胞形成同心圆状排列(图1),而同侧DNLL中细胞未排列形成明显的同心圆状(图2)。对侧DNLL中的HRP数量较多,占三个亚核中HRP细胞总数 的27%,而同侧DNLL中HRP数量较少,只占HRP细胞总数的10%。
, http://www.100md.com
图1(A) 对侧DNLL中的HRP单标细胞 ×40
图2(B) 同侧DNLL及INLL的HRP单标细胞 ×40
INLL和VNLL:仅在同侧的INLL和VNLL中观察到HRP细胞(图2,3),体积较DNLL的为小,以中小圆形、卵圆形或长梭形为主,突起较DNLL的为少,细胞长轴多与外侧丘系纤维平行。INLL的HRP细胞较多,占HRP细胞总数的22%(图2),而VNLL的HRP细胞排列密集,数量最多,约占HRP细胞总数的41%(图3)。
图3(C) 同侧VNLL的HRP单标细胞 ×40
2.2 GABA标记细胞
在低倍镜下,GABA阳性神经元的胞浆呈均匀的棕黄色,在高倍镜下,则可在胞浆中观察到棕黄色的细小颗粒。NLL的三个亚核都有染色强度及数量不等的GABA免疫阳性细胞。其中DNLL中的GABA细胞免疫染色最强,细胞大小不等,形态为多极、梭形、圆形和卵圆形。INLL和VNLL中的 GABA细胞染色较弱,体积较小,为中、小圆形或卵圆形。由于本实验研究的重点在NLL-IC投射细胞,故未对NLL各亚核中GABA细胞进行计数。
, 百拇医药
2.3 HRP/GABA双标细胞
双标记细胞的特点是在均匀的棕黄色背景沉积着蓝黑色的颗粒。在DNLL中(图4),虽然HRP单标细胞形态各异,大小不等,HRP/GABA双标记细胞却以中、小圆形或卵圆形为主,大多极细胞和长梭形细胞却极少被双染。双标细胞数占投射细胞总数的比例,对侧DNLL为75%,同侧DNLL为51%。在INLL和VNLL中也有大量的HRP/GABA双标细胞,以中等圆形或卵圆形为主(图5,6)。双标细胞占投射细胞的比例,INLL为63%,而VNLL为70%。各亚核的双标细胞数占三个亚核总双标细胞数的比例,对侧DNLL为 29%,同侧DNLL为8%,同侧INLL为21%,同侧VNLL为42%。
, http://www.100md.com
图4(D) 对侧DNLL中的HRP单标细胞(
)、GABA单标细胞(↑)和HRP/GABA双标细胞() ×160图5(E) 同侧INLL中的HRP单标细胞(
)、GABA单标细胞(↑)和HRP/GABA双标细胞() ×200图6(F) 同侧VNLL中的HRP单标细胞(
)、GABA单标细胞(↑)和HRP/GABA双标细胞() ×200, http://www.100md.com
3 讨论
3.1 DNLL-IC的GABA能投射
既往的实验表明,DNLL发出纤维支配双侧IC,对侧支配强于同侧[4]。本实验将HRP注入到一侧ICC后,在双侧DNLL观察到大量HRP单标细胞,对侧DNLL内的单标细胞约为同侧的2.7倍,结果与前人的研究一致。有人发现DNLL-IC投射的终末内具有扁平和多形两种囊泡,而这两种囊泡被普遍认为是抑制性突触的特征性结构[5]。Shneiderman[6]将海人酸注入到一侧DNLL后,观察到双侧 IC的GABA释放减少,以对侧为显著。免疫细胞化学实验也显示,DNLL中大部分神经元均为GABA或GAD阳性神经元[1,2]。以上研究均提示DNLL内可能存在着向IC投射的GABA能神经元。本实验用HRP逆行追踪与免疫细胞化学双标记法研究,发现双侧DNLL的许多神经元为HRP/GABA双标神经元,对侧约为同侧的3.6倍,因而证实了DNLL-IC投射神经元中有许多是GABA能神经元,即DNLL中许多神经元是以GABA作为神经递质对IC产生抑制性影响,而且对侧强于同侧,从而为以上实验的推测提供了直接的形态学依据。另外,双标细胞主要为中、小圆形细胞,大多极细胞和长梭形细胞很少被双染,这与De Xingzhang[8]的研究结果一致,提示在DNLL内部,不同形态的神经元可能执行不同的功能。
, http://www.100md.com
关于DNLL-IC的GABA能神经元的机能意义,有些生理学实验进行了研究。用利多卡因阻断NLL-ICC传导后,双侧IC的双音抑制现象有所减弱[9];向DNLL内注入兴奋性氨基酸海人酸或电刺激DNLL可增加对侧ICC的声音诱发抑制反应[9];向一侧DNLL注入兴奋性氨基酸拮抗剂犬尿喹啉酸,对侧ICC的双音抑制作用大为减弱, 而且对两耳声音的时间差及强度差的感受能力也将改变[10]。以上证据表明IC的双音抑制部分由对侧DNLL-IC的 GABA能投射所调节,这种调节可能是通过形成前馈性抑制通路来实现[11]。下丘内这些对双音刺激起反应的神经元,对两耳声音的时间差和强度差反应敏感,而声源定位能力则取决于两耳声音的时间差及强度差 。Kelly[12]用海人酸毁损一侧或两侧DNLL或手术横断Probst连合纤维(为由DNLL投射到对侧DNLL和IC的纤维),可导致声源定位缺失及最小听角上抬,提示交叉的NLL-IC的GABA能投射在提高ICC对双音刺激的敏感性及提高声源定位能力方面起着重要作用。
, 百拇医药
本实验的数据显示在对侧DNLL和同侧DNLL中分别约有75%和50%的投射神经元为双标神经元,这意味着还有少量其它性质的神经元。根据Winer[2]的研究结果,DNLL中没有甘氨酸能神经元,因而DNLL-IC投射为甘氨酸能的可能性不大。而甘氨酸和GABA为中枢神经系统中两种最主要的抑制性递质,因此推测这些非GABA能的投射可能为兴奋性的。Shneiderman[5]发现同侧DNLL-IC投射神经元的轴突终未中有圆形囊泡,这种形态的囊泡被普遍认为含有兴奋性递质;DNLL的神经元细胞内记录显示当电刺激Probst纤维时,可引发兴奋性突触后电位[13]。以上实验均支持DNLL-IC投射中除了大部分是以GABA为递质的抑制性投射以外,少数为兴奋性投射的推测。至于这些兴奋性投射是以哪种物质作为递质,还有待于深入研究。
3.2 INLL-IC和VNLL-IC的GABA能投射
本实验在INLL和VNLL二核仅于同侧观察到大量的HRP单标细胞,其中VNLL的HRP细胞数量为INLL的1.9倍,为同侧DNLL的4倍,为对侧DNLL的1.5 倍。表明此二核发出大量纤维投射至同侧IC,而同侧VNLL是NLL-IC投射的最大来源,符合前人的研究结果。在同侧IVLL和VNLL中还观察到大量的HRP/GABA双标细胞,表明这些核团投射到IC的神经元中有许多为GABA能神经元,其中INLL的双标细胞数占NLL核双标细胞总数的21%,而VNLL为42%,表明同侧VNLL是NLL-IC的GABA能投射的最大来源。INLL中的双标细胞数占HRP细胞数的63%,VNLL则为70%,表明INLL-IC及VNLL-IC投射神经元中也可能有其它性质的神经元存在。Winer[2]的研究结果显示,INLL和VNLL中甘氨酸能神经元的数量比GABA能神经元的多,Saint-Marie[14]根据[3H]标记的甘氨酸作放射自显影研究结果,推测VNLL中GABA能神经元与甘氨酸能神经元共存。故尽管本实验结果显示VNLL-IC和INLL-IC投射神经元中大部分为GABA能,但不排除存在着甘氨酸能的VNLL-IC和INLL-IC投射,也不排除这两种递质共存于同一投射神经元的可能。最后的定论有待于进一步的实验研究,其机能意义也有待进一步探讨。
, 百拇医药
① 基金项目:广西青年科学基金资助课题(No.9631009)。
作者简介:李佩英(1962-),女,1984年毕业于广西医科大学系,获学士学位,1997年在广西医科大学获神经解剖学硕士学位。现任桂林医科学院解剖教研室主任、副教授。
参考文献
1 李佩英,岳毅刚,张德兴,等.树
听觉中枢GABA能神经元及终末的分布. 桂林医学院学报,1996,9(3):229~2322 Winer JA,Larue DT,Pollak GD.GABA and glycine in the central auditory system of the mustache bat:structural substrates for inbibitory neuronal organization.J Comp Nenrol,1995,355:317~353
, http://www.100md.com
3 Douglas L Oliver,Jeffery AW,Gretchen E Beckius,et al. Morphology of GABAergic neurons in the inferior colliculus of the cat.J Comp Neurol,1994,340:27~42
4 Merchan MA,Saldana E,Plaza I.Dorsal nucleus of the lateral lemniscus in the rat :concentric organization and tonotopic projection to the inferior colliculus.J Comp Neurol, 1994,342:259~278
5 Shneiderman A,Oliver DL.EM autoradiographic study of the projections from the dorssal nucleus of the lateral lemniscus:a possible source of inhibitory inputs to the inferior colliculus.J Comp Neuro,1989,286:28~47
, 百拇医药
6 Shneiderman A,Chase MB,Rockwood JM,et al.Evidence for a GABAergic projection from the dorsal nucleus of the lateral lemniscus to the inferior colliculus.J Neurochem, 1993,60:72~82
7 Rye.Stabilization of the tetramethylbenzidine (TMB) reaction product:Application for retrograde and anterograde tracing and combination with immunohistochemistry.J Histochem Cytochem,1984,32(11):1145~1150
8 De Xingzhang,Liang Li,Jack B Kell,et al.GABAergic projections from the lateral lemniscus to the inferior colliculus of the rat.Hear Res,1998,117:1~12
, 百拇医药
9 Faingold CL,Boersma-Anderson CA,Randal ME.Stimulation or blockade of the dorsal nucleus of the lateral lemniscus alters binaural and tonic inhibition in contralateral inferior colliculus neurons.Hear Res,1993,69:98~106
10 Kelly JB,Li L.Two soures of inhibition affecting binaural evoked responses in the rat's inferior colliculus:the dorsal nucleus of the lateral lemniscus and the superior olivary complex.HearRes,1997,104:112~126
, http://www.100md.com 11 Shneiderman A,Oliver DL,Henkel C.Connections of the dorsal nucleus of the lateral lemniscus:an inhitibory parallel pathway in the ascending auditory system? J Comp Neurol,1988,278:188~208
12 Kelly JB,Li L,Van Adel B.Sound Localization after kainic acid lesions of the dorsal nucleus of the lateral lemniscus in the albino rat.Behav Neurosci,1996,110: 1445~1455
13 Wu SH,Kelly JB.In vitro brain slicestudies of the rat's dorsal nucleus of the lateral lemniscus I. Membrane and synaptic response properties.J Neurophysiol,1995,73:780~793
14 Saint Marie RL,Baker RA.Neurotransmitter-specific uptake and retrograde transport of [3H] glycine from the inferior colliculus by ipsilateral projections of the superior olivary complex and nuclei of the lateral lemniscus.Brain Res,1990, 524:244~253
收稿 1999-12-01, 百拇医药