�Ȼ������Դ�����Լ����������Ӱ��
���ߣ������ ��һ�� �ҽ� ��Ƽ �����
��λ�������(����ҽ�ƴ�ѧ��������ѧԺ)������� ��һ�� �ҽ� ��Ƽ(��ҽ�ƴ�ѧԤ��ҽѧϵ���������ң����� 100054)
�ؼ��ʣ��Ȼ������������������
��������ѧ��־000402 ժҪ ��Ŀ�ġ� �о��������(IEG)���Ȼ��������Ի����е����á��������� Ӧ��Fos��Jun����������֯��ѧ�������Ը�ǻע���Ȼ�����(0.6 mg/kg��6.0 mg/kg)�Ĵ���ͬ����c-fos,c-jun����ˮƽ���й۲졣������� ��ͼ�������ͳ�Ƽ�������ʾ������Ⱦ��2 h��������֯Ƥ�㡢����CA3����С�Ե�Fos��Jun��������������[Aa(%)]��ƽ���ҶȺ�����ϸ������(n)3���������������ȣ�������������(P<0.05)����Ⱦ����IEG������ڶ����顣���磺�����顢�ͼ�����Ͷ������ں���CA3����Fos�����ƽ���Ҷȷֱ�Ϊ136��8��149��14��178��15�������ۡ� ��ʾ������ʹIEG��Ϊת¼�������Ӳ������Ȼ��������������Ķ��Թ��̡����������������Եķ��ӻ����ṩ��һ����ʵ�����ݡ������齫IEG��Ϊ�Ȼ��������Լ������۵�ЧӦָ�ꡣ
, http://www.100md.com
Effects of chloro methylmercury on the expression of immediate early genes in rat brain
LI Guo-jun,DENG Yi-fu,CHU Jin-hua
(Department of preventive Medicine,Capital University of Medical Sciences,Beijing 100054,China)
Abstract:��Objective�� To study the role of immediate early genes(IEG) in the neurotoxic mechanism of chloro methylmercury.��Methods�� Expression of Fos and Jun in different brain regions of rats treated with chloro methylmercury(0^6 mg/kg and 6^0 mg/kg)was observed by using immunohistochemical methods.��Results�� The observation and image analysis showed that after rats treated by chloro methylmercury for 2 hours,the expression of Fos and Jun in cortex,CA-3 area of hippocampus and cerebellum of rats was more higher than that of controls (P<0^05).For example,the average gray levels of Fos expression in the CA-3 area of hippocampus were 136��8,149��14,178��15 in groups of high and low doses of chloro methylmercury and controls respectively.��Conclusion�� The results indicated that IEG,as a transcription regular factor,could participate in the process of neurotoxicity induced by chloro methylmercury.The study may provide experimental basis for expounding molecular neurotoxic mechanism of chloro methylmercury and suggest IEG as an effect index of detection and assessment in the neurotoxicology of methylmercury.
, ��Ĵҽҩ
Key words��Chloro methylmercury; Immediate early genes; Neurotoxicity
������һ�־������ԵĻ�����Ⱦ�50����ձ�ˮٶ�¼���ļ�ʮ���У�����ѧ�ߴ���̬ѧ������ѧ�����ﻯѧ�ȷ���Լ��������û��ƽ����˴����о�����������һЩѧ˵[1��2]���������ڷ���ˮƽ�ϼ��������û����Բ�ʮ��������������ڷ���������ѧ�о��У��������ڻ���(immediate-early genes,IEG)��������ϵͳ�������������������������Ҫ���������ս��ܵ���ע����������£�IEG������ϸ�����������ֻ�����Ϣ���ݡ�ѧϰ�ͼ�����������̡���������˺��Դ̼�ʱ��IEG(��Ҫ����c-fos,c-jun)����Ϊ�̼�������Ԫ�ĵڶ���ʹ��Ŀ�Ļ����������Ϣ�����н��������Ϊ����Ԫ��һ�֡�������ʹ��[3]����Ӧ��Fos��Jun������֯��ѧ�����о��Ȼ����������ж�ʱ��������֯�е�����ʹIEG����ĸı䣬Ϊ����̽�ּ������Եķ��ӻ��Ƽ�Σ�ն������ṩʵ�����ݡ�
, http://www.100md.com
1 �����뷽��
1.1 �Լ������� �Ȼ�����(MMC,�ձ��ⴿҩ��ҩ��ʽ����)���ÿ���FOS��JUN���¡����(����Santa Cruz��˾)��SP-Kit(����,����Zymed��˾)��ʯ����Ƭ��(�¹���Leitz 1515��)��
1.2 Ⱦ������������ SD���Խ�����������220��240 g�������Ϊ3�飬ÿ��5��6ֻ��һ���Ը�ǻע�����������Ⱦ�����ͼ����顢������ֱ�ע�����Ϊ0.6��6.0 mg/kg���Ȼ�������Һ��������ע������Ũ��Ϊ0.009 kg/L��NaCl������ʵ��ʱ�価����һ�£�Ⱦ��2 h���ע�̶���ȡ�ġ�
1.3 ��ע�̶� ����Ⱦ��2 h������ͱ�����(60 mg/kg��ip)������������ǻ���бո����������������Ҳ�ܣ��������Ķ�����37 ��������ˮ100 ml���ٳ�ϴ����֮��4 �������Ũ��Ϊ0.04 kg/L�Ķ�ۼ�ȩ(pH7.4)100 ml��ע������0.5��1 h��
, ��Ĵҽҩ
1.4 ȡ����Ƭ ��ע��Ϻ��ͷ��Ѹ��ǯ���ǣ�����Ӳ��Ĥ������Ĥ��ȡ����������֯����ʸ״�淽������н�����֯�г��������룬��4 ���0.04 kg/L��ۼ�ȩ�м����̶�12��24 h���顢��ˮ��ʯ������ʸ״������Ƭ��Ƭ��5 ��m��
1.5 �����黯��Ӧ���� ��Ƭ������ˮ�����������Ϊ0.03��H2O2-�״�(30 min)��������Ũ��Ϊ0.001 kg/L���ȵ���ø(37 �棬15 min)��������Ũ��Ϊ0.02 kg/L��BSA(37 �棬1 h)����Fos(��Jun)һ��(����Ũ��1��200,4 ��)��ҹ���������ػ�����(����Ũ��1��100)��37 �棬20 min����S-A/HRP(����Ũ��1��100)��37 �棬20 min�����������0.01 mol/L PBS(pH7.4)ϴ��3��5 min,DAB��ɫ��PBS��ֹ��Ӧ����ˮ��������Ƭ��
1.6 ����ж� �����Ȳ�λ�ı��ϲ��հ�������������������嶨λͼ�ס�[4]����Fos,Jun���Ե�����������֯��ƬΪ���Զ��գ���������Ѫ�����Fos,Junһ����Ϊ������գ���0.01 mol/LPBS(pH7.4)����Fos,Junһ����Ϊ�հ��ա�
, ��Ĵҽҩ
1.7 ������ ���ձ���OLYMPUS BH1��������ϵͳ��
1.8 ͼ����� ��Mias-2000ͼ�����ϵͳ���������黯��Ӧ��Ƭ���ж���������ÿ����Ƭ�ֱ�Դ���Ƥ�㡢����CA3����С�Թ۲�2����Ұ���ӳ����Ϊ46 151.811 17 ��m2���ֱ�ⶨ���������[Aa(%)]��ƽ���Ҷȼ�����ϸ������(n)�Ȳ�����
1.9 ͳ��ѧ���� ����PEMS Ver.2.1���з��������
2 ���
2.1 �Ȼ������Դ���ͬ����Fos���ױ����Ӱ�� �⾵�¹۲죬Ⱦ�����������֯��Fos����������֯��ѧ��Ӧ���ʳ��ػ�ɫ����״��λ����ϸ�������ֽ���ϸ�����ڣ��������в��ֱ������CA-3��FosȾɫ���Ե���ϸ���ֲ��ܼ�������ʶ���Σ�������ɫ�ĺ˳�Բ�λ���Բ�Σ���ɫ��(ͼ1)��С��FosȾɫ���Ե��ѿ�Ұ��ϸ���ʵ������У��������ɫ����(ͼ2)��Ƥ��������ϸ����ɢ�ڷֲ���Ƥ��IJ�ͬ������ϸ����̬���졣������������Ӧ��λ��������ϸ��Fos���Ա����ɫdz(ͼ3)��������ռ��հ��վ�δ�����Ա��
, ��Ĵҽҩ
��ͼ�����ϵͳ�£��ⶨ����Fos��������������[Aa(%)],ƽ���Ҷȼ�����ϸ������(n)�Ȳ���������1����ͳ��ѧ���飬�ڴ���Ƥ�㣬�ͼ����������Aa(%)��n�����ڶ�����(P<0.05)���ں���CA3�����ͼ�����ƽ���Ҷȼ�������Aa(%)��ƽ���Ҷ��������Ƚϣ�������������(P<0.05)����С����ϸ�������ͼ�����������ƽ���ҶȾ����ڶ�����(P<0.05��P<0.01)��Aa(%)���ڶ�����(P<0.05)�����Ͻ������Ⱦ����Fos������������ǿ���Ͼ����ڶ����顣
��1 �Ȼ�������С��ͬ����Fos���ױ����Ӱ��(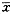 ��s) ���
��s) ���
����Ƥ��
����CA3��
С����ϸ��
, ��Ĵҽҩ
Aa(%)
ƽ���Ҷ�
n
Aa(%)
ƽ���Ҷ�
n
Aa(%)
ƽ���Ҷ�
n
������
8.45��2.55��*
171��2*
, ��Ĵҽҩ
47��8*
15.17��7.83��
136��8��
32��13
4.42��0.50����
160��13��**��
9��2
�ͼ�����
6.66��1.24��
189��8
40��6��
, ��Ĵҽҩ
7.27��3.78
149��14��
23��10
2.95��0.35
180��7*
8��2
������
1.84��1.03
194��12
29��3
3.68��0.93
178��15
, ��Ĵҽҩ
26��5
2.15��0.49
208��5
5��1
�ͼ����飺0.6 mg/kg���Ȼ������� �����飺6.0 mg/kg���Ȼ������� Aa(%)����������ȣ� n������ϸ����
�������Ƚϣ���P<0.05,����P<0.01; ��ͼ�����Ƚϣ���P<0.05��2 �Ȼ������Դ���ͬ����Jun���ױ����Ӱ��(��s) ���
����Ƥ��
����CA3��
, ��Ĵҽҩ
С����ϸ��
Aa(%)
ƽ���Ҷ�
n
Aa(%)
ƽ���Ҷ�
n
Aa(%)
ƽ���Ҷ�
n
������
6.76��4.30
166��6*
, ��Ĵҽҩ
28��1
9.60��2.42��
149��10*
30��9
3.37��0.68
158��8����
8
��
2
�ͼ�����
4.54��1.82
175��14*
, ��Ĵҽҩ
23��2
7.66��2.94��
156��12*
28��11
3.16��1.79
184��9��
9
��
3
������
2.92��1.98
198��12
, http://www.100md.com
26��8
4.42��1.02
174��11
26��8
2.28��0.14
202��2
6
��
1
�������Ƚϣ���P<0.05,����P<0.01;��ͼ�����Ƚϣ���P<0.05
2.2 �Ȼ������Դ���ͬ����Jun���ױ����Ӱ�� �⾵�¹۲죬����Jun�ı����ص�ͬFos(ͼ4��6)����ͳ��ѧ���飬�ڴ���Ƥ�㣬�ͼ�������������ƽ���Ҷ����������ȣ�������������(P<0.05)��������λJun�ı�����ɻ���ͬFos(����2)�����Ͻ������Ⱦ����Jun�ı�����������ǿ���Ͼ����ڶ����顣
, http://www.100md.com
3 ����
3.1 �����Ķ����ü���IEG����Ӱ����ص� �������ڸ������ʣ���֬���ԣ�����Ѫ�����ϻ�̥�����Ͻ�����˻�̥������֯�������Ի����Լ����ж�����1953�����ձ����ֵġ�ˮٶ������60������ҹ��ɻ���������ؽ�����Ҳ���ֹ��̶Ȳ��ȵļ����ж��ı��֡�����������֯����ԭ�������ʹ��ϸ�����������Ա䡣����Ϊ���صIJ�λ��С�Ժʹ����������ٴ��ϱ���ΪС���Թ���ʧ������Ұ��С�����ԡ��о��������ϰ��ȡ�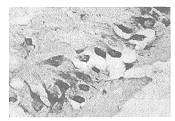
ͼ1 �߹��麣��CA-3��Fos����(��400)
ͼ2 ����С��Fos����(��400)
, ��Ĵҽҩ
ͼ3 �����麣��CA-3��Fos����(��400)
ͼ4 �߹��麣��CA-3��Jun����(��400)
ͼ5 ����С��Jun����(��400)
ͼ6 ������С��Jun����(��400)
��ʵ����ø�ǻע��Ⱦ�����۲��Ȼ������Դ�������֯�������ڻ���(IEG������Fos,Jun��)�����Ӱ�켰�ص㡣�����ʾ��(1)�����ɽϹ㷺�������ڸ���Ҫ�����������Ƥ�㡢��������С�����ȣ���������������������IEG�ı�������С�����ڼ��������£���Щ������Fos,Jun�ı�����������ǿ���Ͼ��������ߣ��Ҿ���ͳ��ѧ����(P<0.05)��(2)������Ҫ�����ڸ���������ϸ���Ͳ��ֽ���ϸ����ʹ��Щϸ�����ڵ�Fos,Jun���׳��ָ߱��(3)���ż���Ⱦ�����������ߣ�Fos��Jun�ı���Ҳ�в�ͬ�̶ȵ����ߣ�������С����������������ͳ��ѧ����(P<0.05)���ڱ�ʵ�������±��ֳ�һ���ļ���-��Ӧ(ЧӦ)��ϵ���������˵���������յ���������֯��c-fos��c-jun����ĵ��ױ��
, ��Ĵҽҩ
3.2 ������IEG����Ӱ��Ŀ��ܹ��̺ͻ��� ��ʵ��۲�Ľ����ʾ���������յ�Fos,Jun�����ʵĺϳɲ�����һ�����������á����е��о�������c-fos��c-jun��ԭ�������е����ּ������(IEG)������������£���ϸ�����е�ˮƽ����������Fos��Jun���������ϸ������Ҫת¼���ӣ�����ͨ�������������γ���Դ�����弤���-1(AP-1)��������Ϊת¼��������������AP-1���λ����֬(TPA)��Ӧ���н�ϣ�Ӱ�����ڰл���ı�������c-fos,c-jun�ļ���ͱ������Ϊ��ͻ����Ĺ��ܱ�־[5]���ڲ���״̬�£����ִ̼���������ʹ����ˮ�����������˵��˺��Դ̼���������������Ԫc-fos,c-jun����ı��c-fos mRNA����߷���1h��Fos���������黯��Ӧ���ֵ��2h[6]��Fos/Jun��Ϊ������ʹ��ͨ������AP-1���λ������˴ӵڶ���ʹ�����ڻ��������ź�ת����
��c-fos��c-jun���յ����ﱾ��Ҳ�����˱Ƚϸ��ӵ�һϵ�м������ữ��Ӧ�����й����ױ��������������ӻ�����ص������£����յ�c-fos��c-jun�����ɡ������Ϊ���Ұ��ἤø�����ֱ��Ӱ��������øC-��1(PLC-��1)�����ữ��һ��PLC-��1���ض��л��������ữ�����ɽ���֬��������������(PIP2)ˮ��������ἡ��(IP3)�Ͷ�����������(DAG)��IP3������ڴ�Ca2+�ͷ����ӣ�ϸ��������Ca2+���࣬���ͬʱ��Ca2+��DAG���ɼ����øC(PKC)[7]��PKC��һ��ʹRaf-1���ữ�����������Raf-1������MAPK-K(mitogen-activated protein kinase-kinase),ʹ֮���ữ[8]�����Ե�MAPK-Kͨ����Erk�Ұ�����հ���л����ữ����ʹ�����ؼ���ĵ���ø(MAPK)�����[9]��MAPK��ʹת¼����p62TCF���ữ����һ�����°�����p62TCF����Ԫ������(TCF)���ߣ����յ�����c-fos��c-jun[8��10]��
, ��Ĵҽҩ
�������ڹ����о����Ѽ�������Ӧ����������ɼ����Ȼ������Ͷ��Ȼ�������DNA�������ã����Ҽ���������ϸ��������Ca2+���ӣ�IP3��������[11]����ϱ�����۲쵽�Ľ�����ɳ����жϣ�����c-fos��c-jun���յ��������ˮƽ�������õ���Ҫ����֮һ��������c-fos��c-jun���յ����̿������������ı��������������յ�c-fos��c-jun���������ƣ�������ϸ�������д��ڽ�һ���о�֤ʵ��
3.3 �����յ�IEG��������弰IEG��Ӧ��ǰ�� c-fos��̬ѧ��ʾ��ͨ������Ϊ��ϵ���ܻ����̬��λָ�꣬���м�ⷽ�㡢�ֱ��ʸ�(�ﵥ��ϸ��ˮƽ)���ŵ�[12]����������δ����������о��������з������������á����о�����IEG���յ�����ȿ�����̽�������������õİ�ϸ�����������ڶ���̽�ּ���-ЧӦ(��Ӧ)��ϵ����ʾ�����ڼ������о��������У�IEG���бȽ���Ҫ�����������DZ�ڵ�Ӧ�ü�ֵ��
, ��Ĵҽҩ
���⣬��Ǧ���Ի��Ƶ��о��У�Ҳ����IEG����ʾǦ���õ��Բ�λ�Ͱ�ϸ������������ȾǦ���������ߣ�IEG�ı���Ҳ����������[13]��c-fos��c-jun�ı�����Ǧ��һ��������ϸ����������Ҫ�ٷ�����֮һ[14]���ɴ˿ɼ���IEG��һЩ�ؽ�������Ķ������ù����У����в��ɺ��ӵ�Ӱ������á����ڶ������ﶾ�������У������־��(biological markers)��ѡ��ͽ����Ǻܹؼ��ģ���ȡ���ڶ������û��Ƶ��о������
���ݱ��о��Ľ����Ŀǰ��֪̽��IEG�յ�����Ļ��ƣ���ʵ���ҽ��飬��IEG��Ϊ�������Լ������۵�ЧӦָ�꣬Ϊ���Լ������������Σ�ն����۵춨һ���Ļ��������������ȡ�����������һ����֤��
�����
[1] �����棬�����.�Ȼ������Դ����Զ�ަ�������Ӱ��.����ҽ�ƴ�ѧѧ����1994��25��388-392.
, ��Ĵҽҩ
[2] Yangama H,Horigome T,Ichimura T,et al.Differential effects of methylmercury on the phosphorylation of protein species in the brain of acutely intoxicated rats.Toxicology,1994,92:101.
[3] Morgan S,Greenberg ME.The regulation and function of c-fos and other immediate early genes in the nervous system.Neuron,1990,4:477-485.
[4] ��������˹����.���������嶨λͼ��.�������������������磬1991.40-135.
[5] �����ţ�����.����������ѧ.�����������ҽ�����磬1995.43.
, ��Ĵҽҩ
[6] MacDonald MC,Robertson HA,Wilkinson M.Age-and dose-related NMDA induction of FOS-like immunoreactivity and c-fos mRNA in the arcuate nucleus of immature female rats.Dev Brain Res,1993,73:193-198.
[7] Berridge MJ.Inositol trisphosphate and calcium signalling.Nature,1993,361:315-325.
[8] Kolch W,Heldecker G,Kochs G,et al.Protein kinase Ca activates RAF-1 by direct phophorylation.Nature,1993,364:249-252.
, ��Ĵҽҩ
[9] Kyriakis JM,App H,Zhang XF,et al.Raf-1 activates MAP kinase-kinase.Nature,1992,358:417-421.
[10] Gille H,Sharrocks AD�� Shaw PE.Phophorylation of transcription factor p62TCF by MAP kinase stimulates ternary complex formation at c-fos promoter.Nature,1992,358:414-417.
[11] Sarafian TA.Methylmercury increase intracellular Ca2+ and inositol phosphate level in cultured cerebellar granule neurons.J Neurochem,1993,61:648.
[12] ������������.��ѧ��Ҫ.����������ҽ�ƴ�ѧ�й�ҽ�ƴ�ѧ���ϳ����磬1993.23-24��272-285.
[13] ��������������Ф��������.Ǧ�Դ���ͬ����IEG�����Ӱ��.�����о���1999��28��65-69.
[14] ����������������.Ǧ��������ϵͳc-Fos,c-Jun��ϸ��������Ӱ���о�.����.����.ͻ�䣬1999��11.322-323.
(�ո����ڣ�2000-01-05), http://www.100md.com
��λ�������(����ҽ�ƴ�ѧ��������ѧԺ)������� ��һ�� �ҽ� ��Ƽ(��ҽ�ƴ�ѧԤ��ҽѧϵ���������ң����� 100054)
�ؼ��ʣ��Ȼ������������������
��������ѧ��־000402 ժҪ ��Ŀ�ġ� �о��������(IEG)���Ȼ��������Ի����е����á��������� Ӧ��Fos��Jun����������֯��ѧ�������Ը�ǻע���Ȼ�����(0.6 mg/kg��6.0 mg/kg)�Ĵ���ͬ����c-fos,c-jun����ˮƽ���й۲졣������� ��ͼ�������ͳ�Ƽ�������ʾ������Ⱦ��2 h��������֯Ƥ�㡢����CA3����С�Ե�Fos��Jun��������������[Aa(%)]��ƽ���ҶȺ�����ϸ������(n)3���������������ȣ�������������(P<0.05)����Ⱦ����IEG������ڶ����顣���磺�����顢�ͼ�����Ͷ������ں���CA3����Fos�����ƽ���Ҷȷֱ�Ϊ136��8��149��14��178��15�������ۡ� ��ʾ������ʹIEG��Ϊת¼�������Ӳ������Ȼ��������������Ķ��Թ��̡����������������Եķ��ӻ����ṩ��һ����ʵ�����ݡ������齫IEG��Ϊ�Ȼ��������Լ������۵�ЧӦָ�ꡣ
, http://www.100md.com
Effects of chloro methylmercury on the expression of immediate early genes in rat brain
LI Guo-jun,DENG Yi-fu,CHU Jin-hua
(Department of preventive Medicine,Capital University of Medical Sciences,Beijing 100054,China)
Abstract:��Objective�� To study the role of immediate early genes(IEG) in the neurotoxic mechanism of chloro methylmercury.��Methods�� Expression of Fos and Jun in different brain regions of rats treated with chloro methylmercury(0^6 mg/kg and 6^0 mg/kg)was observed by using immunohistochemical methods.��Results�� The observation and image analysis showed that after rats treated by chloro methylmercury for 2 hours,the expression of Fos and Jun in cortex,CA-3 area of hippocampus and cerebellum of rats was more higher than that of controls (P<0^05).For example,the average gray levels of Fos expression in the CA-3 area of hippocampus were 136��8,149��14,178��15 in groups of high and low doses of chloro methylmercury and controls respectively.��Conclusion�� The results indicated that IEG,as a transcription regular factor,could participate in the process of neurotoxicity induced by chloro methylmercury.The study may provide experimental basis for expounding molecular neurotoxic mechanism of chloro methylmercury and suggest IEG as an effect index of detection and assessment in the neurotoxicology of methylmercury.
, ��Ĵҽҩ
Key words��Chloro methylmercury; Immediate early genes; Neurotoxicity
������һ�־������ԵĻ�����Ⱦ�50����ձ�ˮٶ�¼���ļ�ʮ���У�����ѧ�ߴ���̬ѧ������ѧ�����ﻯѧ�ȷ���Լ��������û��ƽ����˴����о�����������һЩѧ˵[1��2]���������ڷ���ˮƽ�ϼ��������û����Բ�ʮ��������������ڷ���������ѧ�о��У��������ڻ���(immediate-early genes,IEG)��������ϵͳ�������������������������Ҫ���������ս��ܵ���ע����������£�IEG������ϸ�����������ֻ�����Ϣ���ݡ�ѧϰ�ͼ�����������̡���������˺��Դ̼�ʱ��IEG(��Ҫ����c-fos,c-jun)����Ϊ�̼�������Ԫ�ĵڶ���ʹ��Ŀ�Ļ����������Ϣ�����н��������Ϊ����Ԫ��һ�֡�������ʹ��[3]����Ӧ��Fos��Jun������֯��ѧ�����о��Ȼ����������ж�ʱ��������֯�е�����ʹIEG����ĸı䣬Ϊ����̽�ּ������Եķ��ӻ��Ƽ�Σ�ն������ṩʵ�����ݡ�
, http://www.100md.com
1 �����뷽��
1.1 �Լ������� �Ȼ�����(MMC,�ձ��ⴿҩ��ҩ��ʽ����)���ÿ���FOS��JUN���¡����(����Santa Cruz��˾)��SP-Kit(����,����Zymed��˾)��ʯ����Ƭ��(�¹���Leitz 1515��)��
1.2 Ⱦ������������ SD���Խ�����������220��240 g�������Ϊ3�飬ÿ��5��6ֻ��һ���Ը�ǻע�����������Ⱦ�����ͼ����顢������ֱ�ע�����Ϊ0.6��6.0 mg/kg���Ȼ�������Һ��������ע������Ũ��Ϊ0.009 kg/L��NaCl������ʵ��ʱ�価����һ�£�Ⱦ��2 h���ע�̶���ȡ�ġ�
1.3 ��ע�̶� ����Ⱦ��2 h������ͱ�����(60 mg/kg��ip)������������ǻ���бո����������������Ҳ�ܣ��������Ķ�����37 ��������ˮ100 ml���ٳ�ϴ����֮��4 �������Ũ��Ϊ0.04 kg/L�Ķ�ۼ�ȩ(pH7.4)100 ml��ע������0.5��1 h��
, ��Ĵҽҩ
1.4 ȡ����Ƭ ��ע��Ϻ��ͷ��Ѹ��ǯ���ǣ�����Ӳ��Ĥ������Ĥ��ȡ����������֯����ʸ״�淽������н�����֯�г��������룬��4 ���0.04 kg/L��ۼ�ȩ�м����̶�12��24 h���顢��ˮ��ʯ������ʸ״������Ƭ��Ƭ��5 ��m��
1.5 �����黯��Ӧ���� ��Ƭ������ˮ�����������Ϊ0.03��H2O2-�״�(30 min)��������Ũ��Ϊ0.001 kg/L���ȵ���ø(37 �棬15 min)��������Ũ��Ϊ0.02 kg/L��BSA(37 �棬1 h)����Fos(��Jun)һ��(����Ũ��1��200,4 ��)��ҹ���������ػ�����(����Ũ��1��100)��37 �棬20 min����S-A/HRP(����Ũ��1��100)��37 �棬20 min�����������0.01 mol/L PBS(pH7.4)ϴ��3��5 min,DAB��ɫ��PBS��ֹ��Ӧ����ˮ��������Ƭ��
1.6 ����ж� �����Ȳ�λ�ı��ϲ��հ�������������������嶨λͼ�ס�[4]����Fos,Jun���Ե�����������֯��ƬΪ���Զ��գ���������Ѫ�����Fos,Junһ����Ϊ������գ���0.01 mol/LPBS(pH7.4)����Fos,Junһ����Ϊ�հ��ա�
, ��Ĵҽҩ
1.7 ������ ���ձ���OLYMPUS BH1��������ϵͳ��
1.8 ͼ����� ��Mias-2000ͼ�����ϵͳ���������黯��Ӧ��Ƭ���ж���������ÿ����Ƭ�ֱ�Դ���Ƥ�㡢����CA3����С�Թ۲�2����Ұ���ӳ����Ϊ46 151.811 17 ��m2���ֱ�ⶨ���������[Aa(%)]��ƽ���Ҷȼ�����ϸ������(n)�Ȳ�����
1.9 ͳ��ѧ���� ����PEMS Ver.2.1���з��������
2 ���
2.1 �Ȼ������Դ���ͬ����Fos���ױ����Ӱ�� �⾵�¹۲죬Ⱦ�����������֯��Fos����������֯��ѧ��Ӧ���ʳ��ػ�ɫ����״��λ����ϸ�������ֽ���ϸ�����ڣ��������в��ֱ������CA-3��FosȾɫ���Ե���ϸ���ֲ��ܼ�������ʶ���Σ�������ɫ�ĺ˳�Բ�λ���Բ�Σ���ɫ��(ͼ1)��С��FosȾɫ���Ե��ѿ�Ұ��ϸ���ʵ������У��������ɫ����(ͼ2)��Ƥ��������ϸ����ɢ�ڷֲ���Ƥ��IJ�ͬ������ϸ����̬���졣������������Ӧ��λ��������ϸ��Fos���Ա����ɫdz(ͼ3)��������ռ��հ��վ�δ�����Ա��
, ��Ĵҽҩ
��ͼ�����ϵͳ�£��ⶨ����Fos��������������[Aa(%)],ƽ���Ҷȼ�����ϸ������(n)�Ȳ���������1����ͳ��ѧ���飬�ڴ���Ƥ�㣬�ͼ����������Aa(%)��n�����ڶ�����(P<0.05)���ں���CA3�����ͼ�����ƽ���Ҷȼ�������Aa(%)��ƽ���Ҷ��������Ƚϣ�������������(P<0.05)����С����ϸ�������ͼ�����������ƽ���ҶȾ����ڶ�����(P<0.05��P<0.01)��Aa(%)���ڶ�����(P<0.05)�����Ͻ������Ⱦ����Fos������������ǿ���Ͼ����ڶ����顣
��1 �Ȼ�������С��ͬ����Fos���ױ����Ӱ��(
��s) �������Ƥ��
����CA3��
С����ϸ��
, ��Ĵҽҩ
Aa(%)
ƽ���Ҷ�
n
Aa(%)
ƽ���Ҷ�
n
Aa(%)
ƽ���Ҷ�
n
������
8.45��2.55��*
171��2*
, ��Ĵҽҩ
47��8*
15.17��7.83��
136��8��
32��13
4.42��0.50����
160��13��**��
9��2
�ͼ�����
6.66��1.24��
189��8
40��6��
, ��Ĵҽҩ
7.27��3.78
149��14��
23��10
2.95��0.35
180��7*
8��2
������
1.84��1.03
194��12
29��3
3.68��0.93
178��15
, ��Ĵҽҩ
26��5
2.15��0.49
208��5
5��1
�ͼ����飺0.6 mg/kg���Ȼ������� �����飺6.0 mg/kg���Ȼ������� Aa(%)����������ȣ� n������ϸ����
�������Ƚϣ���P<0.05,����P<0.01; ��ͼ�����Ƚϣ���P<0.05��2 �Ȼ������Դ���ͬ����Jun���ױ����Ӱ��(
��s) �������Ƥ��
����CA3��
, ��Ĵҽҩ
С����ϸ��
Aa(%)
ƽ���Ҷ�
n
Aa(%)
ƽ���Ҷ�
n
Aa(%)
ƽ���Ҷ�
n
������
6.76��4.30
166��6*
, ��Ĵҽҩ
28��1
9.60��2.42��
149��10*
30��9
3.37��0.68
158��8����
8
��
2
�ͼ�����
4.54��1.82
175��14*
, ��Ĵҽҩ
23��2
7.66��2.94��
156��12*
28��11
3.16��1.79
184��9��
9
��
3
������
2.92��1.98
198��12
, http://www.100md.com
26��8
4.42��1.02
174��11
26��8
2.28��0.14
202��2
6
��
1
�������Ƚϣ���P<0.05,����P<0.01;��ͼ�����Ƚϣ���P<0.05
2.2 �Ȼ������Դ���ͬ����Jun���ױ����Ӱ�� �⾵�¹۲죬����Jun�ı����ص�ͬFos(ͼ4��6)����ͳ��ѧ���飬�ڴ���Ƥ�㣬�ͼ�������������ƽ���Ҷ����������ȣ�������������(P<0.05)��������λJun�ı�����ɻ���ͬFos(����2)�����Ͻ������Ⱦ����Jun�ı�����������ǿ���Ͼ����ڶ����顣
, http://www.100md.com
3 ����
3.1 �����Ķ����ü���IEG����Ӱ����ص� �������ڸ������ʣ���֬���ԣ�����Ѫ�����ϻ�̥�����Ͻ�����˻�̥������֯�������Ի����Լ����ж�����1953�����ձ����ֵġ�ˮٶ������60������ҹ��ɻ���������ؽ�����Ҳ���ֹ��̶Ȳ��ȵļ����ж��ı��֡�����������֯����ԭ�������ʹ��ϸ�����������Ա䡣����Ϊ���صIJ�λ��С�Ժʹ����������ٴ��ϱ���ΪС���Թ���ʧ������Ұ��С�����ԡ��о��������ϰ��ȡ�
ͼ1 �߹��麣��CA-3��Fos����(��400)
ͼ2 ����С��Fos����(��400)
, ��Ĵҽҩ
ͼ3 �����麣��CA-3��Fos����(��400)
ͼ4 �߹��麣��CA-3��Jun����(��400)
ͼ5 ����С��Jun����(��400)
ͼ6 ������С��Jun����(��400)
��ʵ����ø�ǻע��Ⱦ�����۲��Ȼ������Դ�������֯�������ڻ���(IEG������Fos,Jun��)�����Ӱ�켰�ص㡣�����ʾ��(1)�����ɽϹ㷺�������ڸ���Ҫ�����������Ƥ�㡢��������С�����ȣ���������������������IEG�ı�������С�����ڼ��������£���Щ������Fos,Jun�ı�����������ǿ���Ͼ��������ߣ��Ҿ���ͳ��ѧ����(P<0.05)��(2)������Ҫ�����ڸ���������ϸ���Ͳ��ֽ���ϸ����ʹ��Щϸ�����ڵ�Fos,Jun���׳��ָ߱��(3)���ż���Ⱦ�����������ߣ�Fos��Jun�ı���Ҳ�в�ͬ�̶ȵ����ߣ�������С����������������ͳ��ѧ����(P<0.05)���ڱ�ʵ�������±��ֳ�һ���ļ���-��Ӧ(ЧӦ)��ϵ���������˵���������յ���������֯��c-fos��c-jun����ĵ��ױ��
, ��Ĵҽҩ
3.2 ������IEG����Ӱ��Ŀ��ܹ��̺ͻ��� ��ʵ��۲�Ľ����ʾ���������յ�Fos,Jun�����ʵĺϳɲ�����һ�����������á����е��о�������c-fos��c-jun��ԭ�������е����ּ������(IEG)������������£���ϸ�����е�ˮƽ����������Fos��Jun���������ϸ������Ҫת¼���ӣ�����ͨ�������������γ���Դ�����弤���-1(AP-1)��������Ϊת¼��������������AP-1���λ����֬(TPA)��Ӧ���н�ϣ�Ӱ�����ڰл���ı�������c-fos,c-jun�ļ���ͱ������Ϊ��ͻ����Ĺ��ܱ�־[5]���ڲ���״̬�£����ִ̼���������ʹ����ˮ�����������˵��˺��Դ̼���������������Ԫc-fos,c-jun����ı��c-fos mRNA����߷���1h��Fos���������黯��Ӧ���ֵ��2h[6]��Fos/Jun��Ϊ������ʹ��ͨ������AP-1���λ������˴ӵڶ���ʹ�����ڻ��������ź�ת����
��c-fos��c-jun���յ����ﱾ��Ҳ�����˱Ƚϸ��ӵ�һϵ�м������ữ��Ӧ�����й����ױ��������������ӻ�����ص������£����յ�c-fos��c-jun�����ɡ������Ϊ���Ұ��ἤø�����ֱ��Ӱ��������øC-��1(PLC-��1)�����ữ��һ��PLC-��1���ض��л��������ữ�����ɽ���֬��������������(PIP2)ˮ��������ἡ��(IP3)�Ͷ�����������(DAG)��IP3������ڴ�Ca2+�ͷ����ӣ�ϸ��������Ca2+���࣬���ͬʱ��Ca2+��DAG���ɼ����øC(PKC)[7]��PKC��һ��ʹRaf-1���ữ�����������Raf-1������MAPK-K(mitogen-activated protein kinase-kinase),ʹ֮���ữ[8]�����Ե�MAPK-Kͨ����Erk�Ұ�����հ���л����ữ����ʹ�����ؼ���ĵ���ø(MAPK)�����[9]��MAPK��ʹת¼����p62TCF���ữ����һ�����°�����p62TCF����Ԫ������(TCF)���ߣ����յ�����c-fos��c-jun[8��10]��
, ��Ĵҽҩ
�������ڹ����о����Ѽ�������Ӧ����������ɼ����Ȼ������Ͷ��Ȼ�������DNA�������ã����Ҽ���������ϸ��������Ca2+���ӣ�IP3��������[11]����ϱ�����۲쵽�Ľ�����ɳ����жϣ�����c-fos��c-jun���յ��������ˮƽ�������õ���Ҫ����֮һ��������c-fos��c-jun���յ����̿������������ı��������������յ�c-fos��c-jun���������ƣ�������ϸ�������д��ڽ�һ���о�֤ʵ��
3.3 �����յ�IEG��������弰IEG��Ӧ��ǰ�� c-fos��̬ѧ��ʾ��ͨ������Ϊ��ϵ���ܻ����̬��λָ�꣬���м�ⷽ�㡢�ֱ��ʸ�(�ﵥ��ϸ��ˮƽ)���ŵ�[12]����������δ����������о��������з������������á����о�����IEG���յ�����ȿ�����̽�������������õİ�ϸ�����������ڶ���̽�ּ���-ЧӦ(��Ӧ)��ϵ����ʾ�����ڼ������о��������У�IEG���бȽ���Ҫ�����������DZ�ڵ�Ӧ�ü�ֵ��
, ��Ĵҽҩ
���⣬��Ǧ���Ի��Ƶ��о��У�Ҳ����IEG����ʾǦ���õ��Բ�λ�Ͱ�ϸ������������ȾǦ���������ߣ�IEG�ı���Ҳ����������[13]��c-fos��c-jun�ı�����Ǧ��һ��������ϸ����������Ҫ�ٷ�����֮һ[14]���ɴ˿ɼ���IEG��һЩ�ؽ�������Ķ������ù����У����в��ɺ��ӵ�Ӱ������á����ڶ������ﶾ�������У������־��(biological markers)��ѡ��ͽ����Ǻܹؼ��ģ���ȡ���ڶ������û��Ƶ��о������
���ݱ��о��Ľ����Ŀǰ��֪̽��IEG�յ�����Ļ��ƣ���ʵ���ҽ��飬��IEG��Ϊ�������Լ������۵�ЧӦָ�꣬Ϊ���Լ������������Σ�ն����۵춨һ���Ļ��������������ȡ�����������һ����֤��
�����
[1] �����棬�����.�Ȼ������Դ����Զ�ަ�������Ӱ��.����ҽ�ƴ�ѧѧ����1994��25��388-392.
, ��Ĵҽҩ
[2] Yangama H,Horigome T,Ichimura T,et al.Differential effects of methylmercury on the phosphorylation of protein species in the brain of acutely intoxicated rats.Toxicology,1994,92:101.
[3] Morgan S,Greenberg ME.The regulation and function of c-fos and other immediate early genes in the nervous system.Neuron,1990,4:477-485.
[4] ��������˹����.���������嶨λͼ��.�������������������磬1991.40-135.
[5] �����ţ�����.����������ѧ.�����������ҽ�����磬1995.43.
, ��Ĵҽҩ
[6] MacDonald MC,Robertson HA,Wilkinson M.Age-and dose-related NMDA induction of FOS-like immunoreactivity and c-fos mRNA in the arcuate nucleus of immature female rats.Dev Brain Res,1993,73:193-198.
[7] Berridge MJ.Inositol trisphosphate and calcium signalling.Nature,1993,361:315-325.
[8] Kolch W,Heldecker G,Kochs G,et al.Protein kinase Ca activates RAF-1 by direct phophorylation.Nature,1993,364:249-252.
, ��Ĵҽҩ
[9] Kyriakis JM,App H,Zhang XF,et al.Raf-1 activates MAP kinase-kinase.Nature,1992,358:417-421.
[10] Gille H,Sharrocks AD�� Shaw PE.Phophorylation of transcription factor p62TCF by MAP kinase stimulates ternary complex formation at c-fos promoter.Nature,1992,358:414-417.
[11] Sarafian TA.Methylmercury increase intracellular Ca2+ and inositol phosphate level in cultured cerebellar granule neurons.J Neurochem,1993,61:648.
[12] ������������.��ѧ��Ҫ.����������ҽ�ƴ�ѧ�й�ҽ�ƴ�ѧ���ϳ����磬1993.23-24��272-285.
[13] ��������������Ф��������.Ǧ�Դ���ͬ����IEG�����Ӱ��.�����о���1999��28��65-69.
[14] ����������������.Ǧ��������ϵͳc-Fos,c-Jun��ϸ��������Ӱ���о�.����.����.ͻ�䣬1999��11.322-323.
(�ո����ڣ�2000-01-05), http://www.100md.com