阿托品、荷包牡丹碱对家鸽(Columba livia)顶盖Ⅱa-f亚层与Ⅲ层神经元间突触传递的影响
http://www.100md.com
37c医学网
阿托品、荷包牡丹碱对家鸽(Columba livia)顶盖Ⅱa-f亚层与Ⅲ层神经元间突触传递的影响
余小平 王彬
摘 要 目的:探讨阿托品(Atropine,ATR)、荷包牡丹碱(Bicuculline,BIC)对家鸽顶盖Ⅱa-f亚层与Ⅲ层神经元间突触传递的影响。方法:在孵育离体顶盖脑片标本上,采用脉冲方波电刺激Ⅱa-f亚层,利用玻璃微电极胞外记录技术记录Ⅲ层神经元放电频率。结果:Ⅲ层神经元对电刺激Ⅱa-f亚层有反应的28个单位中,13个单位(占46.4%)表现为增频,15个单位(占53.6%)表现为减频。表现为增频的13个Ⅲ层神经元中10个单位(占76.9%)的增频效应可被ATR部分或完全逆转,另3个单位的阻断效果不明显;15个被Ⅱa-f亚层刺激传入减频的Ⅲ层神经元,其中11个单位(占73.3%)的减频作用可被BIC阻断,另4个单位则不产生阻遏作用。上述药物的作用具有可逆性和重复性。结论:乙酰胆碱(Acetylcholine,ACh)可能参与了家鸽顶盖Ⅱa-f亚层与Ⅲ层神经元间兴奋性突触传递,而γ-氨基丁酸(Gamma-aminobutyric,GABA)则参与其抑制性突触传递。
, http://www.100md.com
关键词:顶盖;脑片;阿托品;荷包牡丹碱
具有高度层状结构的顶盖可分为6层,以Ⅰ~Ⅵ表示,其Ⅱ层又可分为10个亚层,以a~f表示,Ⅱa~f亚层为视网膜主要投射区[1]。资料表明,ACh、GABA在鸽顶盖内含量较高且具高亲摄取特性[2]。家鸽顶盖神经元接收和加工处理视觉信息过程中,ACh主要是与毒蕈碱(Muscarine,M)型受体结合而进行兴奋传递的[3],GABA是与GABAA型受体结合进行抑制性传递[1]。目前有关家鸽顶盖Ⅱa~f亚层与Ⅲ层神经元间的突触传递尚未见报道。本工作利用脑片标本的优点,探讨了ACh的M型3种受体亚型无选择性的经典拮抗挤ATR、以及GABA的GABAA型受体竞争性拮抗挤BIC对顶盖Ⅱa~f亚层与Ⅲ层神经元间突触传递的影响。
, 百拇医药
1 材料与方法
1.1 顶盖脑片的制备
本实验选用280~450g体重的家鸽,雌雄不拘。腹腔注射10%氨基甲酸乙酯(1.2ml/100g体重)后,断头,迅速剪除头部皮肤和颅骨,移去硬脑膜,取出全脑,置于4℃预先充混合气(95%O2,5%CO2)饱和的人工脑脊液(ACSF)中降温片刻,然后分离出一侧顶盖,按冠状面切成500μm厚的脑片。用广口吸管吸取1~3片完整脑片移入人工浴槽中孵育。
1.2 脑片孵育方法
人工浴槽分外室和内室。外室盛有循环流动的蒸馏水,由WC/09型超级恒温仪控制其温度在(37±1)°C,同时通入300~500ml/min流量的混合气体(95%O2,5%CO2)。混合气体经外室加温湿润后扩散入内室,作用于脑片表面。内室容积较小,上面紧贴一层尼龙网,脑片置于尼龙网上,半浸没式孵育。ACSF通过乳胶管流经外室恒温加热后抵达内室,滴速为15~30滴/min,其成分(mmol/L):NaCl124,KCl5,MgSO41.3,KH2PO41.25,NaHCO326,CaCl22.5,葡萄糖10,pH值用磷酸缓冲液(pH5.8)调至7.40±0.05。脑片孵育60~90min后开始记录。
, 百拇医药
1.3 刺激
选用国泰JJC-4型刺激器。刺激频率为100Hz、波宽0.1ms、空载电压5~15V组成的3个方波串刺激。刺激电极为直径0.1mm绝缘不锈钢丝制成的双极电极,两极间距小于0.1mm。
1.4 记录
采用充满2%滂胺天蓝(pH7.4)的醋酸钠导电溶液的玻璃电极作记录电极,尖端直径0.1μm,直流阻抗5~15MΩ。经PF-5型微电极操纵器推送记录电极至Ⅲ层内,记录Ⅲ层神经元的自发放电。实验完毕后,部分脑片用滂胺天蓝标记,然后参考Karten和Hodes的鸽脑图谱[4]作组织学鉴定。检测到的电信号经JZ-802型微电极阻抗变换器和MEZ-8201型微电极放大器放大后显示在SR-76型双线示波器上,并用SZF-10生物电放大器监听放电情况。放大信号同时输入Doctor-852电生理实验智能仪作序列密度直方图并存盘。
, 百拇医药
实验数据经统计学处理后用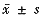 表示。灌流给药前后放电频率用配对t检验。
表示。灌流给药前后放电频率用配对t检验。
2 结果
2.1 电刺激Ⅱa~f亚层对顶盖Ⅲ层神经元自发放电活动的影响,见表1
表1 刺激Ⅱa~f亚层对Ⅲ层神经元电活动的影响
Tab1 The effects of stimulating sublayer Ⅱa~f upon layer Ⅲ neurons
Excitatory
, 百拇医药
Inhibitory
Total
Number
13
15
28
Centigrade(%)
46.4
53.6
100.0
在记录到36个Ⅲ层神经元自发放电基础上,用方波串脉冲电刺激(频率100Hz、波宽0.1ms、空载电压5~15V)顶盖Ⅱa~f亚层,观察对Ⅲ层神经元自发放电的影响。结果显示,28个神经元(28/36,77.8%)在刺激后1~3s内有反应,表现为刺激后放电频率出现短暂的变化。其中,13个单位(13/28,占46.4%)产生兴奋效应,放电频率由刺激前(3.61±2.45)个/s增至(25.65±16.65)个/s,持续时程(2.32±0.93)s,P<0.001,见参考文献[5];15个单位(15/28,占53.6%)则由刺激前(4.12±1.87)个/s减至(0.69±0.53)个/s,持续时程(2.86±1.17)s,P<0.05,见参考文献[6]。差异均具有显著性。
, 百拇医药
2.2 ATR对Ⅱa~f亚层与Ⅲ层神经元间兴奋性突触传递的影响
电刺激(频率100Hz、波宽0.1ms、空载电压5~15V)顶盖Ⅱa~f亚层对Ⅲ层神经元自发放电产生兴奋效应的13例中,观察灌流ATR(10-6mol/L)对刺激效应的影响。结果显示,ATR能阻断电刺激Ⅱa~f亚层对Ⅲ层神经元的增频效应,用方波串刺激Ⅱa~f亚层时,其中10例(占76.9%)对电刺激Ⅱa~f亚层不再产生放电增频现象,平均放电频率仍保持在(3.61±2.45)个/s(P<0.05),去除ATR后,这些神经元能再次被Ⅱa~f亚层刺激传入引起放电增频,见参考文献[5]。另3例(占23.1%)对刺激Ⅱa~f亚层的放电增频反应不受ATR影响。
2.3 BIC对顶盖Ⅱa~f亚层与Ⅲ层神经元间抑制性突触传递的影响
, 百拇医药
电刺激(频率100Hz、波宽0.1ms、空载电压5~15V)Ⅱa~f亚层对Ⅲ层神经元自发放电活动产生抑制性反应的15个单位,灌流10-6mol/LBIC,观察对刺激效应的影响。结果表明,11个单位(占73.3%)对电刺激Ⅱa~f亚层产生的抑制效应能被BIC阻断,表现为刺激时其放电频率不变。经正常ACSF冲洗后,这些单位能再次被电刺激Ⅱa~f亚层所抑制,见参考文献[6]。另4个单位(占26.7%)的刺激抑制效应不受BIC影响,其放电频率仍保持在(4.12±1.87)(P>0.05)。
3 讨论
3.1 顶盖Ⅱa~f亚层与Ⅲ层神经元间的兴奋性突触传递
有关家鸽视网膜、同侧Wulst视区、对侧顶盖及峡核(NI)到顶盖的传入研究较多[7],这些研究提示ACh在其中可能参与了其间的兴奋性传递。但是,顶盖内Ⅱa~f亚层与Ⅲ层神经元间的突触传递,尚未见报道。本实验电刺激Ⅱa~f亚层时,观察到部分Ⅲ层神经元的放电频率增加,且ATR能阻遏这种增频效应,提示Ⅱa~f亚层有兴奋性纤维到达Ⅲ层。刺激Wulst视区和对侧顶盖能使顶盖神经元产生2~17ms潜伏期的兴奋性突触后电位(EPSP),暗示有单或多突触参与了这种EPSP[7]。电刺激视神经可使81%顶盖细胞产生兴奋性突触后电位,经荧光黄(LY)标记显示部分细胞为位于Ⅲ层内的节细胞,从EPSP的潜伏期和频率跟随能力推测视神经末梢与顶盖Ⅲ层神经元间形成多突触兴奋性传递[8]。最近,利用免疫学方法证实峡核-顶盖通路中的兴奋性突触传递是以ACh为递质,作用受体为M型受体[9]。鉴于Wulst视区、对侧顶盖、视网膜及峡核到顶盖的投射区域一致,即Ⅱa~f亚层[9,10],结合本实验的结果,可以推测,这些视结构可能通过ACh纤维传至Ⅱa~f亚层,与Ⅱa~f亚层神经元形成兴奋性突触,Ⅱa~f亚层神经元再通过ACh能纤维传至顶盖Ⅲ层。
, 百拇医药
本实验结果表明顶盖Ⅱa~f亚层与Ⅲ层神经元间的兴奋性突触传递的递质可能是ACh,并通过M型受体起作用,从而进一步证实了ACh在顶盖内起着兴奋性递质的作用以及M型受体在顶盖内广泛分布这一理论。
3.2 顶盖Ⅱa~f亚层与Ⅲ层神经元之间的抑制性突触传递
实验中观察到电刺激Ⅱa~f亚层时,放电频率发生显著变化的28个Ⅲ层神经元中,有15个单位(占53.6%)的放电频率减少,应用BIC后能阻断这种刺激效应,表明GABA可能参与了Ⅱa~f亚层与Ⅲ层神经元间的抑制性突触传递。
据在体实验研究的文献报道,去除峡核可引起顶盖脱抑制。刺激峡核小细胞部可使顶盖Ⅲ层神经元产生单或多突触IPSP(Inhibitory postsynapse potential),这种抑制性突触后电位部分是通过GABA与GABAA型受体结合起作用的[7,11]。刺激顶盖表面及Wulst视区可以通过GABA能中间神经元使Ⅲ层神经元的电活动受到抑制,这种抑制反应可被BIC阻断[7]。这些资料有力地支持了本实验所得结论。
, http://www.100md.com
基金项目:国家自然科学基金资助项目(39170108)
作者简介:余小平(1970.10),男,重庆市开县人,硕士,助教,主要从事鸟类生理学方面的研究,发表论文5篇。电话:(023)68752291
余小平(第三军医大学预防医学系营养卫生学教研室,重庆 400038)
王彬(西南师范大学生理学教研室,重庆 400715)
参考文献
[1]Toggenburger G,Felix D,Cuenod M,et al.In vitro release of endogenous β-alanine,GABA and glutamate and eletrophysiolo-gical effect of β-alanine in pigeon optic tectum[J].J Neurochem,1982,39(1):176-183.
, http://www.100md.com
[2]Henke H,Schenker T M,Cuenod M.Effect of retinal ablation on uptake of glutamate,glycine,GABA,proline and choline in pigeon tectum[J].J Neurochem,1976,26(1):131-134.
[3]Wang S R,Li Z,Xu H Y.Muscarinic action of acetylcholine in the pigeon optic tectum[J].Exp Neurol,1986,94(2):436-440.
[4]Karten H T,Hodos W A.A stereotaxic atlas of the brain of the pigeon (Columba livia)[M].Baltimore:Johns Hopkins Press,1976.174-193.
, 百拇医药
[5]余小平,王彬.乙酰胆碱对家鸽(ColumbaLivia)顶盖脑片中Ⅲ层神经元电活动的影响[J].重庆师范学院学报(自然科学版),1998,15(3):53-58.
[6]余小平,王彬.γ-氨基丁酸对家鸽(ColumbaLivia)顶盖脑片中Ⅲ层神经元电活动的影响[J].四川师范大学学报(自然科学版),1999,22(1):108-113.
[7]Hardy O,Leresche N,Jassik Gerschen F D.Postsynaptic potentials in neurons of the pigeon optic tectum in response to afferent stimulation from the retina and other visual structure:an intracellular study[J].Brain Res,1984,311(1):65-74.
[8]高宏峰,吴冈义,王书荣.家鸽峡核顶盖通路的神经递质[J].生物物理学报,1995,11(2):233-237.
[9]王荫亭,许红艳,王书荣.家鸽圆核和峡核的免疫细胞化学分析[J].生物物理学报,1995,11(1):43-48.
[10]王荫亭,王书荣.用羰花青荧光染料追踪家鸽顶盖与中脑核团的神经连接[J].生物物理学报,1990,6(1):112-115.
[11]吴冈义,王书荣,颜坤.家鸽顶盖细胞的突触后电位和形态特征[J].中国科学(B辑),1992,20(9):937-943., 百拇医药
余小平 王彬
摘 要 目的:探讨阿托品(Atropine,ATR)、荷包牡丹碱(Bicuculline,BIC)对家鸽顶盖Ⅱa-f亚层与Ⅲ层神经元间突触传递的影响。方法:在孵育离体顶盖脑片标本上,采用脉冲方波电刺激Ⅱa-f亚层,利用玻璃微电极胞外记录技术记录Ⅲ层神经元放电频率。结果:Ⅲ层神经元对电刺激Ⅱa-f亚层有反应的28个单位中,13个单位(占46.4%)表现为增频,15个单位(占53.6%)表现为减频。表现为增频的13个Ⅲ层神经元中10个单位(占76.9%)的增频效应可被ATR部分或完全逆转,另3个单位的阻断效果不明显;15个被Ⅱa-f亚层刺激传入减频的Ⅲ层神经元,其中11个单位(占73.3%)的减频作用可被BIC阻断,另4个单位则不产生阻遏作用。上述药物的作用具有可逆性和重复性。结论:乙酰胆碱(Acetylcholine,ACh)可能参与了家鸽顶盖Ⅱa-f亚层与Ⅲ层神经元间兴奋性突触传递,而γ-氨基丁酸(Gamma-aminobutyric,GABA)则参与其抑制性突触传递。
, http://www.100md.com
关键词:顶盖;脑片;阿托品;荷包牡丹碱
具有高度层状结构的顶盖可分为6层,以Ⅰ~Ⅵ表示,其Ⅱ层又可分为10个亚层,以a~f表示,Ⅱa~f亚层为视网膜主要投射区[1]。资料表明,ACh、GABA在鸽顶盖内含量较高且具高亲摄取特性[2]。家鸽顶盖神经元接收和加工处理视觉信息过程中,ACh主要是与毒蕈碱(Muscarine,M)型受体结合而进行兴奋传递的[3],GABA是与GABAA型受体结合进行抑制性传递[1]。目前有关家鸽顶盖Ⅱa~f亚层与Ⅲ层神经元间的突触传递尚未见报道。本工作利用脑片标本的优点,探讨了ACh的M型3种受体亚型无选择性的经典拮抗挤ATR、以及GABA的GABAA型受体竞争性拮抗挤BIC对顶盖Ⅱa~f亚层与Ⅲ层神经元间突触传递的影响。
, 百拇医药
1 材料与方法
1.1 顶盖脑片的制备
本实验选用280~450g体重的家鸽,雌雄不拘。腹腔注射10%氨基甲酸乙酯(1.2ml/100g体重)后,断头,迅速剪除头部皮肤和颅骨,移去硬脑膜,取出全脑,置于4℃预先充混合气(95%O2,5%CO2)饱和的人工脑脊液(ACSF)中降温片刻,然后分离出一侧顶盖,按冠状面切成500μm厚的脑片。用广口吸管吸取1~3片完整脑片移入人工浴槽中孵育。
1.2 脑片孵育方法
人工浴槽分外室和内室。外室盛有循环流动的蒸馏水,由WC/09型超级恒温仪控制其温度在(37±1)°C,同时通入300~500ml/min流量的混合气体(95%O2,5%CO2)。混合气体经外室加温湿润后扩散入内室,作用于脑片表面。内室容积较小,上面紧贴一层尼龙网,脑片置于尼龙网上,半浸没式孵育。ACSF通过乳胶管流经外室恒温加热后抵达内室,滴速为15~30滴/min,其成分(mmol/L):NaCl124,KCl5,MgSO41.3,KH2PO41.25,NaHCO326,CaCl22.5,葡萄糖10,pH值用磷酸缓冲液(pH5.8)调至7.40±0.05。脑片孵育60~90min后开始记录。
, 百拇医药
1.3 刺激
选用国泰JJC-4型刺激器。刺激频率为100Hz、波宽0.1ms、空载电压5~15V组成的3个方波串刺激。刺激电极为直径0.1mm绝缘不锈钢丝制成的双极电极,两极间距小于0.1mm。
1.4 记录
采用充满2%滂胺天蓝(pH7.4)的醋酸钠导电溶液的玻璃电极作记录电极,尖端直径0.1μm,直流阻抗5~15MΩ。经PF-5型微电极操纵器推送记录电极至Ⅲ层内,记录Ⅲ层神经元的自发放电。实验完毕后,部分脑片用滂胺天蓝标记,然后参考Karten和Hodes的鸽脑图谱[4]作组织学鉴定。检测到的电信号经JZ-802型微电极阻抗变换器和MEZ-8201型微电极放大器放大后显示在SR-76型双线示波器上,并用SZF-10生物电放大器监听放电情况。放大信号同时输入Doctor-852电生理实验智能仪作序列密度直方图并存盘。
, 百拇医药
实验数据经统计学处理后用
2 结果
2.1 电刺激Ⅱa~f亚层对顶盖Ⅲ层神经元自发放电活动的影响,见表1
表1 刺激Ⅱa~f亚层对Ⅲ层神经元电活动的影响
Tab1 The effects of stimulating sublayer Ⅱa~f upon layer Ⅲ neurons
Excitatory
, 百拇医药
Inhibitory
Total
Number
13
15
28
Centigrade(%)
46.4
53.6
100.0
在记录到36个Ⅲ层神经元自发放电基础上,用方波串脉冲电刺激(频率100Hz、波宽0.1ms、空载电压5~15V)顶盖Ⅱa~f亚层,观察对Ⅲ层神经元自发放电的影响。结果显示,28个神经元(28/36,77.8%)在刺激后1~3s内有反应,表现为刺激后放电频率出现短暂的变化。其中,13个单位(13/28,占46.4%)产生兴奋效应,放电频率由刺激前(3.61±2.45)个/s增至(25.65±16.65)个/s,持续时程(2.32±0.93)s,P<0.001,见参考文献[5];15个单位(15/28,占53.6%)则由刺激前(4.12±1.87)个/s减至(0.69±0.53)个/s,持续时程(2.86±1.17)s,P<0.05,见参考文献[6]。差异均具有显著性。
, 百拇医药
2.2 ATR对Ⅱa~f亚层与Ⅲ层神经元间兴奋性突触传递的影响
电刺激(频率100Hz、波宽0.1ms、空载电压5~15V)顶盖Ⅱa~f亚层对Ⅲ层神经元自发放电产生兴奋效应的13例中,观察灌流ATR(10-6mol/L)对刺激效应的影响。结果显示,ATR能阻断电刺激Ⅱa~f亚层对Ⅲ层神经元的增频效应,用方波串刺激Ⅱa~f亚层时,其中10例(占76.9%)对电刺激Ⅱa~f亚层不再产生放电增频现象,平均放电频率仍保持在(3.61±2.45)个/s(P<0.05),去除ATR后,这些神经元能再次被Ⅱa~f亚层刺激传入引起放电增频,见参考文献[5]。另3例(占23.1%)对刺激Ⅱa~f亚层的放电增频反应不受ATR影响。
2.3 BIC对顶盖Ⅱa~f亚层与Ⅲ层神经元间抑制性突触传递的影响
, 百拇医药
电刺激(频率100Hz、波宽0.1ms、空载电压5~15V)Ⅱa~f亚层对Ⅲ层神经元自发放电活动产生抑制性反应的15个单位,灌流10-6mol/LBIC,观察对刺激效应的影响。结果表明,11个单位(占73.3%)对电刺激Ⅱa~f亚层产生的抑制效应能被BIC阻断,表现为刺激时其放电频率不变。经正常ACSF冲洗后,这些单位能再次被电刺激Ⅱa~f亚层所抑制,见参考文献[6]。另4个单位(占26.7%)的刺激抑制效应不受BIC影响,其放电频率仍保持在(4.12±1.87)(P>0.05)。
3 讨论
3.1 顶盖Ⅱa~f亚层与Ⅲ层神经元间的兴奋性突触传递
有关家鸽视网膜、同侧Wulst视区、对侧顶盖及峡核(NI)到顶盖的传入研究较多[7],这些研究提示ACh在其中可能参与了其间的兴奋性传递。但是,顶盖内Ⅱa~f亚层与Ⅲ层神经元间的突触传递,尚未见报道。本实验电刺激Ⅱa~f亚层时,观察到部分Ⅲ层神经元的放电频率增加,且ATR能阻遏这种增频效应,提示Ⅱa~f亚层有兴奋性纤维到达Ⅲ层。刺激Wulst视区和对侧顶盖能使顶盖神经元产生2~17ms潜伏期的兴奋性突触后电位(EPSP),暗示有单或多突触参与了这种EPSP[7]。电刺激视神经可使81%顶盖细胞产生兴奋性突触后电位,经荧光黄(LY)标记显示部分细胞为位于Ⅲ层内的节细胞,从EPSP的潜伏期和频率跟随能力推测视神经末梢与顶盖Ⅲ层神经元间形成多突触兴奋性传递[8]。最近,利用免疫学方法证实峡核-顶盖通路中的兴奋性突触传递是以ACh为递质,作用受体为M型受体[9]。鉴于Wulst视区、对侧顶盖、视网膜及峡核到顶盖的投射区域一致,即Ⅱa~f亚层[9,10],结合本实验的结果,可以推测,这些视结构可能通过ACh纤维传至Ⅱa~f亚层,与Ⅱa~f亚层神经元形成兴奋性突触,Ⅱa~f亚层神经元再通过ACh能纤维传至顶盖Ⅲ层。
, 百拇医药
本实验结果表明顶盖Ⅱa~f亚层与Ⅲ层神经元间的兴奋性突触传递的递质可能是ACh,并通过M型受体起作用,从而进一步证实了ACh在顶盖内起着兴奋性递质的作用以及M型受体在顶盖内广泛分布这一理论。
3.2 顶盖Ⅱa~f亚层与Ⅲ层神经元之间的抑制性突触传递
实验中观察到电刺激Ⅱa~f亚层时,放电频率发生显著变化的28个Ⅲ层神经元中,有15个单位(占53.6%)的放电频率减少,应用BIC后能阻断这种刺激效应,表明GABA可能参与了Ⅱa~f亚层与Ⅲ层神经元间的抑制性突触传递。
据在体实验研究的文献报道,去除峡核可引起顶盖脱抑制。刺激峡核小细胞部可使顶盖Ⅲ层神经元产生单或多突触IPSP(Inhibitory postsynapse potential),这种抑制性突触后电位部分是通过GABA与GABAA型受体结合起作用的[7,11]。刺激顶盖表面及Wulst视区可以通过GABA能中间神经元使Ⅲ层神经元的电活动受到抑制,这种抑制反应可被BIC阻断[7]。这些资料有力地支持了本实验所得结论。
, http://www.100md.com
基金项目:国家自然科学基金资助项目(39170108)
作者简介:余小平(1970.10),男,重庆市开县人,硕士,助教,主要从事鸟类生理学方面的研究,发表论文5篇。电话:(023)68752291
余小平(第三军医大学预防医学系营养卫生学教研室,重庆 400038)
王彬(西南师范大学生理学教研室,重庆 400715)
参考文献
[1]Toggenburger G,Felix D,Cuenod M,et al.In vitro release of endogenous β-alanine,GABA and glutamate and eletrophysiolo-gical effect of β-alanine in pigeon optic tectum[J].J Neurochem,1982,39(1):176-183.
, http://www.100md.com
[2]Henke H,Schenker T M,Cuenod M.Effect of retinal ablation on uptake of glutamate,glycine,GABA,proline and choline in pigeon tectum[J].J Neurochem,1976,26(1):131-134.
[3]Wang S R,Li Z,Xu H Y.Muscarinic action of acetylcholine in the pigeon optic tectum[J].Exp Neurol,1986,94(2):436-440.
[4]Karten H T,Hodos W A.A stereotaxic atlas of the brain of the pigeon (Columba livia)[M].Baltimore:Johns Hopkins Press,1976.174-193.
, 百拇医药
[5]余小平,王彬.乙酰胆碱对家鸽(ColumbaLivia)顶盖脑片中Ⅲ层神经元电活动的影响[J].重庆师范学院学报(自然科学版),1998,15(3):53-58.
[6]余小平,王彬.γ-氨基丁酸对家鸽(ColumbaLivia)顶盖脑片中Ⅲ层神经元电活动的影响[J].四川师范大学学报(自然科学版),1999,22(1):108-113.
[7]Hardy O,Leresche N,Jassik Gerschen F D.Postsynaptic potentials in neurons of the pigeon optic tectum in response to afferent stimulation from the retina and other visual structure:an intracellular study[J].Brain Res,1984,311(1):65-74.
[8]高宏峰,吴冈义,王书荣.家鸽峡核顶盖通路的神经递质[J].生物物理学报,1995,11(2):233-237.
[9]王荫亭,许红艳,王书荣.家鸽圆核和峡核的免疫细胞化学分析[J].生物物理学报,1995,11(1):43-48.
[10]王荫亭,王书荣.用羰花青荧光染料追踪家鸽顶盖与中脑核团的神经连接[J].生物物理学报,1990,6(1):112-115.
[11]吴冈义,王书荣,颜坤.家鸽顶盖细胞的突触后电位和形态特征[J].中国科学(B辑),1992,20(9):937-943., 百拇医药