水稻主基因-多基因混合遗传控制白叶枯病抗性的基因效应分析
作者:王建设 王建康 朱立宏 盖钧镒
单位:王建设(北京市农林科学院蔬菜研究中心,北京 100081);王建康(河南省农业科学院,郑州 450002);朱立宏 盖钧镒(南京农业大学,江苏南京 210095)
关键词:水稻;白叶枯病;主基因-多基因抗性
遗传学报000106
摘要:利用主基因-多基因混合遗传模型分析了5个抗感交组合对水稻白叶枯病菌抗性的基因效应,结果表明5个组合中的3个在主基因抗性遗传符合孟德尔分离比的前提下存在多基因抗性,而且这3个组合彼此间抗病基因的加性效应、主基因和多基因遗传方差及其遗传率存在变异。说明水稻白叶枯病抗性虽以主基因作用为主,但考虑到抗性的持久性,建议在水稻白叶枯病育种中构建主基因-多基因混合遗传体系,以有效抑制白叶枯病菌群体中的小种的波动。
, http://www.100md.com
中图分类号:Q311 文献标识码:A
文章编号:0379-4172(2000)01-0034-0038
Major-Polygene Effect Analysis of Resistance to Bacterial Blight (Xanthomonas campestris pv.oryzae) in Rice
WANG Jian-She
(Beijing Agricultural and Forest Academy,Beijing Vegetable Research Center,Beijing 100081,China)
WANG Jian-Kang
(Henan Agricultural Academy,Zhenzhou 450002,China)
, http://www.100md.com
ZHU Li-Hong,GAI Jun-Yi
(Department of Agronomy,Nangjing Agricultural University,Nangjing 210095,China)
Abstract:Five crosses between resistance and susceptible were analyzed to study major-polygene effect using major-polygene mixed mode.The result showed that 3 of 5 crosses were controlled by both major gene and polygene.In addition,there were large variation of additive effect,variance as well as heredity of major gene polygene in 3 crosses.Major gene was predominant in resistant variation,but durability should be considered.We suggested that construct major-polygene system be constructed in the long run in breeding program to ensure a durable and high level of resistance to constrain fluctuation of races of Xanthomonas campestris pv.oryzae population.
, http://www.100md.com
Key words:rice;bacterial blight;major-polygene effect analysis of resistance
杂交稻对白叶枯病菌的抗性主要受加性效应基因控制,但同时也受非加性基因效应的影响,表现加性效应的基因既存在主基因,也存在微效多基因[1]。本文试图证明在主基因抗性符合孟德尔分离比的前提下,鉴定微效多基因抗性的有无及大小,以评价微效多基因在水稻抗白叶枯病育种中的应用前景,为杂交稻亲本选配提供理论依据。
1 材料和方法
1.1 实验材料与杂交设计
8072为组合TN1/DV85杂交后代,携带显性抗病基因X[2,3]a-7。TN1、桂朝2号、IR24、双桂1号和珍珠矮为对白叶枯病菌系ZHE-173感病。利用8072与5个感病品种配制的抗感交组合的亲本、F1及F2群体分析水稻主基因-多基因混合遗传控制抗性的基因效应.
, http://www.100md.com
1.2 遗传分析模型
借助王建康等[4]提出的主基因-多基因混合遗传模型分析主基因-多基因混合遗传控制抗性的基因效应。
1.2.1 主基因-多基因混合遗传模型的建立 将F2世代个体表现型分解为:X=m+g+c+e,其中
m为群体均值,g为主基因效应,c为多基因效应值,e为环境效应。并假定主基因与多基因之间不存在互作,基因型效应与环境效应之间独立。主基因效应为一固定值,多基因效应c~N(0,σ2pg)是随机变量,σ2pg为多基因效应的方差。环境变异e~N(0,σ2e)是随机变量,σ2e为环境方差。若主基因有K个不同的基因型值,则F2群体表现为K个正态分布的混合,通过对混合分布的分解将主基因效应分解出来。除主基因之外的变异是多基因变异和环境变异的混合,记σ2=σ2pg+σ2e。
, 百拇医药
1.2.2 主基因遗传效应的估计 表现完全显性主基因(A-a)其F2群体分布分解为2个成分分布,且2个成分分布具有3:1的孟德尔分离比,其个体主基因基因型分别为Aa、AA和aa,其分布均值μ1和μ2分别为:μ1=m-d,μ2=m+d;m和d极大似然估计分别为: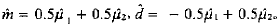
 和
和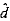 的方差分别为:
的方差分别为: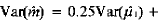
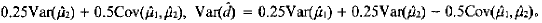 主基因遗传方差和遗传率分别为:σ2mg=0.75d2,h2mg=σ2mg/(σ2mg+σ2)=3d2/(3d2+4σ2)。
主基因遗传方差和遗传率分别为:σ2mg=0.75d2,h2mg=σ2mg/(σ2mg+σ2)=3d2/(3d2+4σ2)。
, http://www.100md.com
1.2.3 多基因存在的鉴定 P1,P2,F1和F2群体的一些统计参数表1所示,样本似然函数:L(ψ)=∏f1(X1i;μ1,σ2e)∏f2(X2i;μ2,σ2e)∏f3(X3i;μ3,σ2e)∏p(Xi;ψ)构建零假设Ho:σ2=σ2e(不存在多基因)和备择假设Ho:σ2>σ2e(有多基因存在,且σ2pg=σ2=σ2e)通过计算两种假设下似然函数的最大值Lo和La,构造似然比统计量:X=2(logLa-logLo)~X2(1)进而对上述假设进行显著性测验。
, http://www.100md.com
表1 P1、P2、F1、F2群体的一些统计参数
Table1 Parameter of P1、P2、F1、F2 population 群体
Population
样本量
Sample xize
样本值
Sample value
均值
Mean
, 百拇医药
方差
Variance
群体分布特征
Population distribution
概率密度函数
Density function
P1
n1
x1i
μ1
σ2e
, 百拇医药
单一正态分布
normal distribution
f1(μ1,σ2e)
P2
n2
x2i
μ2
σ2e
单一正态分布
normal distribution
, 百拇医药
f2(μ1,σ2e)
F1
n3
x3i
μ3
σ2e
单一正态分布
normal distribution
f3(μ1,σ2e)
, http://www.100md.com
F2
n4
x4i
正态混合分布
normal mixture
p(ψ)*
2 结果分析
8072与TN1、桂朝2号、IR24、双桂1号和珍珠矮配制的5个抗感交组合的F2群体经X2测验,结果表明5个组合抗感遗传符合孟德尔分离比(3:1)(表2),说明它们的抗性由显性抗病基因Xa-7控制。
, http://www.100md.com
表2 8072抗性的遗传分析
Table2 Genetic analysis of resistance source 8072 组合
Combination
R
S
x2(3:1)
P
TN1/8072
161
48
0.1330
, 百拇医药
0.75
桂朝2号/8072
(Guichao 2/8072)
124
54
2.1363
0.14
IR24/8072
183
63
0.0135
0.91
, 百拇医药
双桂1号/8072
(Shuanggui1/8072)
123
50
1.2486
0.26
珍珠矮/8072
(Zhengzhuai/8072)
129
44
0.0044
0.95
, http://www.100md.com
利用主基因-多基因混合遗传模型首先确定F2群体的分布均包含2个成分分布(表3),也证明存在一个主基因抗性位点Xa-7,并表现为完全显性,同时同一组合的2个成分分布以及不同组合的同一成分分布的权重存在差异。其次鉴定5个抗感交组合发现其中3个(TN1/8072、桂朝2号/8072和珍珠矮/8072)存在多基因抗性(表4),并估计了抗性基因的加性效应、主基因和多基因的遗传方差及其遗传率(表5)。从表5看出,这3个组合抗性基因的加性效应分别为5.06、5.67和7.390;主基因的遗传方差及其遗传率分别为19.2、24.11和49.96及58.16%、68.59%和73.87%;多基因的遗传方差及其遗传率分别为8.61、9.18和9.36及26.08%、26.12%和16.88%;主基因-多基因遗传率分别为84.24%、94.71%和90.75%。这3个组合抗病基因的加性效应、主基因和多基因遗传方差及其遗传率的变异表明:(1)遗传背景不同的3个感病品种(TN1、桂朝2号和珍珠矮)彼此间与抗性有关的多基因加性效应的大小存在差异,解释了这3个感病品种彼此间感病度高低的数量差异,同时也表明品种感病与抗病的相对性,即品种表现感病,并不是说它完全没有携带抗病基因,只是它携带的抗病基因效应较小,品种表现抗病基因效应较大;(2)8072携带的主效抗基因Xa-7在不同遗传背景中的表达存在差异,暗示着不同遗传背景存在影响Xa-7表达的修饰基;(3)在主基因-多基因混合遗传控制的抗性中,抗性的遗传变异以主基因影响为主,多基因影响为辅.表2 F2混合分布中各参数的极大似然估计
, 百拇医药
Table3 Maximum likelihood estimates in F2 population 组合
Combination
基本分布
Component No.
权重(π)
Weight(π)
均值(μ)
Mean(μ)
方差(σ2)
Variance(σ2)
, 百拇医药
TN1/8072
抗病植株(R-plant)
0.76
5.03
5.20
感病植株(S-plant)
0.24
17.15
5.20
桂朝2号/8072
(Gui chao2/8072)
抗病植株(R-plant)
, 百拇医药
0.70
4.00
7.48
感病植株(S-plant)
0.30
15.34
7.48
IR24/8072
抗病植株(R-plant)
0.75
3.26
5.45
, http://www.100md.com
感病植株(S-plant)
0.25
15.07
5.45
双桂1号/8072
(Shuang gui1/8072)
抗病植株(R-plant)
0.71
4.05
5.49
感病植株(S-plant)
0.29
, http://www.100md.com
15.08
5.49
表4 极大似然法鉴定抗性多基因
Table4 Identification of resistant polygene using maximum likelihood 组合
Combination
maxLa
MaxLo
λ
P
TN1/8072
-710.02
, 百拇医药
-713.03
6.02
<0.05
桂朝2号/8072
(Guichao 2/8072)
-583.13
-591.83
17.40
<0.05
IR24/8072
-752.67
-753.38
, 百拇医药
1.44
>0.05
双桂1号/8072
(Shuanggui1/8072)
-573.82
-574.26
0.88
>0.05
珍珠矮/8072
(Zhengzhuai/8072)
-646.63
-653.13
, http://www.100md.com
12.99
<0.05
表5 主基因-多基因遗传达室效应和遗传方差估计
Table5 Maximum likelihood estimates of major gene effects and genetic variance of major gene 组合
Combination
加性效应
Additive
effect
(d)
主基因
, 百拇医药
遗传方差
Variance of
major gene
(σ2mg)
主基因
遗传率
Heritability of
major gene
(h2mg)%
多基因
遗传方差
, 百拇医药
Variance of
polygene
(σ2pg)
多基因
遗传率
Heritability of
polygene
(h2pg)%
主基因-多
基因遗传率
Heriability of
, http://www.100md.com
major-minor gene
(h2g)%
TN1/8072
5.06
19.20
58.16
8.16
26.08
84.24
桂朝2号/8072
(Guichao 2/8072)
, 百拇医药
5.67
24.11
68.59
9.18
26.12
94.71
珍珠矮/8072
(Zhengzhuai/8072)
7.39
40.96
73.87
9.36
, http://www.100md.com 16.88
90.76
3 讨论
杂交稻对白叶枯病菌的抗性主要受加性效应基因控制,但同时也受非加性基因效应的影响,表现加性效应的基因既存在主基因,也存在微效多基因[1]。利用王建康等[6]提出的主基因-多基因混合遗传模型,分析了5个抗感交组合,结果发现这5个组合在主基因抗性遗传符合孟德尔分离比的前提下,其中3个组合存在多基因抗性,而且这3个组合抗病基因的加性效应、主基因和多基因遗传方差及其遗传的存在变异。说明抗感划分标准是相对的,因菌系、品种、组合而异[5,10]。
前文视分析抗性配合力的固定模型为随机模型,估计了不育系杂种接种菌系ZHE-173时抗性的遗传率,估计的h2B和h2N分别为95.93%和82.67%[1]。本文利用主基因-多基因混合遗传模分析3个组合主基因-多基因遗传率变幅为84.24~94.71%,基中主基因引起F2单株抗性变幅为58.16~73.87%,多基因引起F2单株抗性变幅为16.88~26.08%。一方面说明抗性主要受基因加性效应影响,显性效应和上位性效应影响较小,暗示杂交稻亲本选配中,最好选择基因加性效应大或一般配合力效应高的不育系与恢复系配制杂交组合,预期获得高水平的抗性;另一方面说明在抗病育种中,抗病基因的利用以主基因为主,但考虑到抗性的持久性,应该有意识地积累高水平的多基因控制数量抗性,构建主基因-多基因混合性遗传体系,以有效抑制病原物群体中小种的波动.[7~10]。
, 百拇医药
参考文献
[1]王建设.杂交水稻抗白叶枯病的遗传与抗病基因Xa-7的RFLP标记定位.博士学位论文,南京农业大学,1996.
[2]高山林,朱立宏.水稻白叶枯病抗性遗传研究.南京农学院学报,1982,1:22~23.
[3]朱立宏,万建明,等.籼稻抗白叶枯病衍生抗原选育及遗传分析.见:熊振明.梁承邺主编水稻育种技术基础研究论文集,中国科学技术出版社,1991,24~30.
[4]王建康等.利用F2世代鉴定数量性状主基因-多基因混合遗传模型并估计其遗传效应.遗传学报,1997,24(5):332~440.
[5]章 琦.我国水稻白叶枯病抗性遗传的评价和利用.中国农业科学,1991,24(3):26~36.
, http://www.100md.com
[6]Kock M F.Aspects of quantitative resistance to Xanthomonas campestris pv.oryzae in rice.Ph.D Thesis Agriculture University Wageningen,1990.
[7]Mew T W.Current status and future prospects of research on bacterial blight of rice.Ann.Rev.Phytopatho,1987,125: 539~582.
[8]Mew T W.Changes in race frequency of Xanthomonas oryzae pv.oryzae in response for rice cultivars planted in the phillippines.Plant Disease,1992,76:1029~1032.
, http://www.100md.com
[9]Yamada T.Studies on genetics and breeding of resistance to bacterial leaf blight in rice (VI) Inheritance of quantitative resistance of the variety IR28 to bacterial groups Ⅱ,Ⅲ,Ⅵ of Xanthomonas campestris pv.oryzae of Japan.Japanese Journal Breeding,1984,34:181~190.
[10]Yoshimura A.Genetic of bacterial blight resistance a breeding line of rice.Phytopathology,1984,74:773~777., http://www.100md.com
单位:王建设(北京市农林科学院蔬菜研究中心,北京 100081);王建康(河南省农业科学院,郑州 450002);朱立宏 盖钧镒(南京农业大学,江苏南京 210095)
关键词:水稻;白叶枯病;主基因-多基因抗性
遗传学报000106
摘要:利用主基因-多基因混合遗传模型分析了5个抗感交组合对水稻白叶枯病菌抗性的基因效应,结果表明5个组合中的3个在主基因抗性遗传符合孟德尔分离比的前提下存在多基因抗性,而且这3个组合彼此间抗病基因的加性效应、主基因和多基因遗传方差及其遗传率存在变异。说明水稻白叶枯病抗性虽以主基因作用为主,但考虑到抗性的持久性,建议在水稻白叶枯病育种中构建主基因-多基因混合遗传体系,以有效抑制白叶枯病菌群体中的小种的波动。
, http://www.100md.com
中图分类号:Q311 文献标识码:A
文章编号:0379-4172(2000)01-0034-0038
Major-Polygene Effect Analysis of Resistance to Bacterial Blight (Xanthomonas campestris pv.oryzae) in Rice
WANG Jian-She
(Beijing Agricultural and Forest Academy,Beijing Vegetable Research Center,Beijing 100081,China)
WANG Jian-Kang
(Henan Agricultural Academy,Zhenzhou 450002,China)
, http://www.100md.com
ZHU Li-Hong,GAI Jun-Yi
(Department of Agronomy,Nangjing Agricultural University,Nangjing 210095,China)
Abstract:Five crosses between resistance and susceptible were analyzed to study major-polygene effect using major-polygene mixed mode.The result showed that 3 of 5 crosses were controlled by both major gene and polygene.In addition,there were large variation of additive effect,variance as well as heredity of major gene polygene in 3 crosses.Major gene was predominant in resistant variation,but durability should be considered.We suggested that construct major-polygene system be constructed in the long run in breeding program to ensure a durable and high level of resistance to constrain fluctuation of races of Xanthomonas campestris pv.oryzae population.
, http://www.100md.com
Key words:rice;bacterial blight;major-polygene effect analysis of resistance
杂交稻对白叶枯病菌的抗性主要受加性效应基因控制,但同时也受非加性基因效应的影响,表现加性效应的基因既存在主基因,也存在微效多基因[1]。本文试图证明在主基因抗性符合孟德尔分离比的前提下,鉴定微效多基因抗性的有无及大小,以评价微效多基因在水稻抗白叶枯病育种中的应用前景,为杂交稻亲本选配提供理论依据。
1 材料和方法
1.1 实验材料与杂交设计
8072为组合TN1/DV85杂交后代,携带显性抗病基因X[2,3]a-7。TN1、桂朝2号、IR24、双桂1号和珍珠矮为对白叶枯病菌系ZHE-173感病。利用8072与5个感病品种配制的抗感交组合的亲本、F1及F2群体分析水稻主基因-多基因混合遗传控制抗性的基因效应.
, http://www.100md.com
1.2 遗传分析模型
借助王建康等[4]提出的主基因-多基因混合遗传模型分析主基因-多基因混合遗传控制抗性的基因效应。
1.2.1 主基因-多基因混合遗传模型的建立 将F2世代个体表现型分解为:X=m+g+c+e,其中
m为群体均值,g为主基因效应,c为多基因效应值,e为环境效应。并假定主基因与多基因之间不存在互作,基因型效应与环境效应之间独立。主基因效应为一固定值,多基因效应c~N(0,σ2pg)是随机变量,σ2pg为多基因效应的方差。环境变异e~N(0,σ2e)是随机变量,σ2e为环境方差。若主基因有K个不同的基因型值,则F2群体表现为K个正态分布的混合,通过对混合分布的分解将主基因效应分解出来。除主基因之外的变异是多基因变异和环境变异的混合,记σ2=σ2pg+σ2e。
, 百拇医药
1.2.2 主基因遗传效应的估计 表现完全显性主基因(A-a)其F2群体分布分解为2个成分分布,且2个成分分布具有3:1的孟德尔分离比,其个体主基因基因型分别为Aa、AA和aa,其分布均值μ1和μ2分别为:μ1=m-d,μ2=m+d;m和d极大似然估计分别为:
和的方差分别为:主基因遗传方差和遗传率分别为:σ2mg=0.75d2,h2mg=σ2mg/(σ2mg+σ2)=3d2/(3d2+4σ2)。, http://www.100md.com
1.2.3 多基因存在的鉴定 P1,P2,F1和F2群体的一些统计参数表1所示,样本似然函数:L(ψ)=∏f1(X1i;μ1,σ2e)∏f2(X2i;μ2,σ2e)∏f3(X3i;μ3,σ2e)∏p(Xi;ψ)构建零假设Ho:σ2=σ2e(不存在多基因)和备择假设Ho:σ2>σ2e(有多基因存在,且σ2pg=σ2=σ2e)通过计算两种假设下似然函数的最大值Lo和La,构造似然比统计量:X=2(logLa-logLo)~X2(1)进而对上述假设进行显著性测验。
, http://www.100md.com
表1 P1、P2、F1、F2群体的一些统计参数
Table1 Parameter of P1、P2、F1、F2 population 群体
Population
样本量
Sample xize
样本值
Sample value
均值
Mean
, 百拇医药
方差
Variance
群体分布特征
Population distribution
概率密度函数
Density function
P1
n1
x1i
μ1
σ2e
, 百拇医药
单一正态分布
normal distribution
f1(μ1,σ2e)
P2
n2
x2i
μ2
σ2e
单一正态分布
normal distribution
, 百拇医药
f2(μ1,σ2e)
F1
n3
x3i
μ3
σ2e
单一正态分布
normal distribution
f3(μ1,σ2e)
, http://www.100md.com
F2
n4
x4i
正态混合分布
normal mixture
p(ψ)*
2 结果分析
8072与TN1、桂朝2号、IR24、双桂1号和珍珠矮配制的5个抗感交组合的F2群体经X2测验,结果表明5个组合抗感遗传符合孟德尔分离比(3:1)(表2),说明它们的抗性由显性抗病基因Xa-7控制。
, http://www.100md.com
表2 8072抗性的遗传分析
Table2 Genetic analysis of resistance source 8072 组合
Combination
R
S
x2(3:1)
P
TN1/8072
161
48
0.1330
, 百拇医药
0.75
桂朝2号/8072
(Guichao 2/8072)
124
54
2.1363
0.14
IR24/8072
183
63
0.0135
0.91
, 百拇医药
双桂1号/8072
(Shuanggui1/8072)
123
50
1.2486
0.26
珍珠矮/8072
(Zhengzhuai/8072)
129
44
0.0044
0.95
, http://www.100md.com
利用主基因-多基因混合遗传模型首先确定F2群体的分布均包含2个成分分布(表3),也证明存在一个主基因抗性位点Xa-7,并表现为完全显性,同时同一组合的2个成分分布以及不同组合的同一成分分布的权重存在差异。其次鉴定5个抗感交组合发现其中3个(TN1/8072、桂朝2号/8072和珍珠矮/8072)存在多基因抗性(表4),并估计了抗性基因的加性效应、主基因和多基因的遗传方差及其遗传率(表5)。从表5看出,这3个组合抗性基因的加性效应分别为5.06、5.67和7.390;主基因的遗传方差及其遗传率分别为19.2、24.11和49.96及58.16%、68.59%和73.87%;多基因的遗传方差及其遗传率分别为8.61、9.18和9.36及26.08%、26.12%和16.88%;主基因-多基因遗传率分别为84.24%、94.71%和90.75%。这3个组合抗病基因的加性效应、主基因和多基因遗传方差及其遗传率的变异表明:(1)遗传背景不同的3个感病品种(TN1、桂朝2号和珍珠矮)彼此间与抗性有关的多基因加性效应的大小存在差异,解释了这3个感病品种彼此间感病度高低的数量差异,同时也表明品种感病与抗病的相对性,即品种表现感病,并不是说它完全没有携带抗病基因,只是它携带的抗病基因效应较小,品种表现抗病基因效应较大;(2)8072携带的主效抗基因Xa-7在不同遗传背景中的表达存在差异,暗示着不同遗传背景存在影响Xa-7表达的修饰基;(3)在主基因-多基因混合遗传控制的抗性中,抗性的遗传变异以主基因影响为主,多基因影响为辅.表2 F2混合分布中各参数的极大似然估计
, 百拇医药
Table3 Maximum likelihood estimates in F2 population 组合
Combination
基本分布
Component No.
权重(π)
Weight(π)
均值(μ)
Mean(μ)
方差(σ2)
Variance(σ2)
, 百拇医药
TN1/8072
抗病植株(R-plant)
0.76
5.03
5.20
感病植株(S-plant)
0.24
17.15
5.20
桂朝2号/8072
(Gui chao2/8072)
抗病植株(R-plant)
, 百拇医药
0.70
4.00
7.48
感病植株(S-plant)
0.30
15.34
7.48
IR24/8072
抗病植株(R-plant)
0.75
3.26
5.45
, http://www.100md.com
感病植株(S-plant)
0.25
15.07
5.45
双桂1号/8072
(Shuang gui1/8072)
抗病植株(R-plant)
0.71
4.05
5.49
感病植株(S-plant)
0.29
, http://www.100md.com
15.08
5.49
表4 极大似然法鉴定抗性多基因
Table4 Identification of resistant polygene using maximum likelihood 组合
Combination
maxLa
MaxLo
λ
P
TN1/8072
-710.02
, 百拇医药
-713.03
6.02
<0.05
桂朝2号/8072
(Guichao 2/8072)
-583.13
-591.83
17.40
<0.05
IR24/8072
-752.67
-753.38
, 百拇医药
1.44
>0.05
双桂1号/8072
(Shuanggui1/8072)
-573.82
-574.26
0.88
>0.05
珍珠矮/8072
(Zhengzhuai/8072)
-646.63
-653.13
, http://www.100md.com
12.99
<0.05
表5 主基因-多基因遗传达室效应和遗传方差估计
Table5 Maximum likelihood estimates of major gene effects and genetic variance of major gene 组合
Combination
加性效应
Additive
effect
(d)
主基因
, 百拇医药
遗传方差
Variance of
major gene
(σ2mg)
主基因
遗传率
Heritability of
major gene
(h2mg)%
多基因
遗传方差
, 百拇医药
Variance of
polygene
(σ2pg)
多基因
遗传率
Heritability of
polygene
(h2pg)%
主基因-多
基因遗传率
Heriability of
, http://www.100md.com
major-minor gene
(h2g)%
TN1/8072
5.06
19.20
58.16
8.16
26.08
84.24
桂朝2号/8072
(Guichao 2/8072)
, 百拇医药
5.67
24.11
68.59
9.18
26.12
94.71
珍珠矮/8072
(Zhengzhuai/8072)
7.39
40.96
73.87
9.36
, http://www.100md.com 16.88
90.76
3 讨论
杂交稻对白叶枯病菌的抗性主要受加性效应基因控制,但同时也受非加性基因效应的影响,表现加性效应的基因既存在主基因,也存在微效多基因[1]。利用王建康等[6]提出的主基因-多基因混合遗传模型,分析了5个抗感交组合,结果发现这5个组合在主基因抗性遗传符合孟德尔分离比的前提下,其中3个组合存在多基因抗性,而且这3个组合抗病基因的加性效应、主基因和多基因遗传方差及其遗传的存在变异。说明抗感划分标准是相对的,因菌系、品种、组合而异[5,10]。
前文视分析抗性配合力的固定模型为随机模型,估计了不育系杂种接种菌系ZHE-173时抗性的遗传率,估计的h2B和h2N分别为95.93%和82.67%[1]。本文利用主基因-多基因混合遗传模分析3个组合主基因-多基因遗传率变幅为84.24~94.71%,基中主基因引起F2单株抗性变幅为58.16~73.87%,多基因引起F2单株抗性变幅为16.88~26.08%。一方面说明抗性主要受基因加性效应影响,显性效应和上位性效应影响较小,暗示杂交稻亲本选配中,最好选择基因加性效应大或一般配合力效应高的不育系与恢复系配制杂交组合,预期获得高水平的抗性;另一方面说明在抗病育种中,抗病基因的利用以主基因为主,但考虑到抗性的持久性,应该有意识地积累高水平的多基因控制数量抗性,构建主基因-多基因混合性遗传体系,以有效抑制病原物群体中小种的波动.[7~10]。
, 百拇医药
参考文献
[1]王建设.杂交水稻抗白叶枯病的遗传与抗病基因Xa-7的RFLP标记定位.博士学位论文,南京农业大学,1996.
[2]高山林,朱立宏.水稻白叶枯病抗性遗传研究.南京农学院学报,1982,1:22~23.
[3]朱立宏,万建明,等.籼稻抗白叶枯病衍生抗原选育及遗传分析.见:熊振明.梁承邺主编水稻育种技术基础研究论文集,中国科学技术出版社,1991,24~30.
[4]王建康等.利用F2世代鉴定数量性状主基因-多基因混合遗传模型并估计其遗传效应.遗传学报,1997,24(5):332~440.
[5]章 琦.我国水稻白叶枯病抗性遗传的评价和利用.中国农业科学,1991,24(3):26~36.
, http://www.100md.com
[6]Kock M F.Aspects of quantitative resistance to Xanthomonas campestris pv.oryzae in rice.Ph.D Thesis Agriculture University Wageningen,1990.
[7]Mew T W.Current status and future prospects of research on bacterial blight of rice.Ann.Rev.Phytopatho,1987,125: 539~582.
[8]Mew T W.Changes in race frequency of Xanthomonas oryzae pv.oryzae in response for rice cultivars planted in the phillippines.Plant Disease,1992,76:1029~1032.
, http://www.100md.com
[9]Yamada T.Studies on genetics and breeding of resistance to bacterial leaf blight in rice (VI) Inheritance of quantitative resistance of the variety IR28 to bacterial groups Ⅱ,Ⅲ,Ⅵ of Xanthomonas campestris pv.oryzae of Japan.Japanese Journal Breeding,1984,34:181~190.
[10]Yoshimura A.Genetic of bacterial blight resistance a breeding line of rice.Phytopathology,1984,74:773~777., http://www.100md.com