普通小麦与东方旱麦草异附加系和异代换系的选育与原位杂交检测
作者:张桂芳 刘建文 黄远樟 丁敏 唐顺学 贾旭
单位:张桂芳 刘建文 黄远樟 丁敏(北京师范大学生命科学学院,北京 100875);唐顺学 贾旭(中国科学院遗传研究所,北京 100101)
关键词:旱麦草;普通小麦;附加系;代换系;基因组原位杂交(GISH)
遗传学报000109
摘要:旱麦草属(Eremopyrum)是用于小麦品种改良的又一潜在的植物资源。为了筛选小麦-旱麦草异附加系、异代换系,对普通小麦品种Fukuho×东方旱麦草属间杂种的BC2F3代材料的96粒种子进行了染色体数目的检测,共检出15粒2n=43的种子,8粒2n=44的种子,进一步对以上材料进行的基因组DNA原位杂交,共鉴定出3个单体附加系,2个二体附加系,1个双单体附加,1个小麦三体单体附加,1个附加3条东方旱麦草染色体的小麦单体,在染色体数为42的个体中,检测出1个单体代换,1个双单体代换。根据BC2F3代自交品系来源的不同,初步认为由双单体附加自交比单体附加自交选择异附加系的效率高。
, http://www.100md.com
中图分类号:Q343 文献标识码:A
文章编号:0379-4172(2000)01-0050-0055
Isolation and Verification of Triticum aestivum-Eremopyrum orientale Addition Lines and Substitution Lines by GISH
ZHANG Gui-Fang,LIU Jian-Wen,HUANG Yuan-Zhang,DING Min
(College of Life Sciences,Beijing Normal University,Beijing 100875,China)
TANG Shun-Xue,JIA Xu
, http://www.100md.com
(Institute of Genetics,Chinese Academy of Sciences,Beijing 100101,China)
Abstract:Eremopyrum is a potential genus in wheat improvement.To breed T.aestivum-Er.orientale additional and substitutional lines,chromosome number of 96 BC2F3 individuals were accounted.Fifteen plants with 2n=43 and eight plants with 2n=44 were checked out.GISH results of the individuals with 43 chromosomes showed that they blonged to three different monosomic additions.Two disomic additions,one double-monosomic addition,one trible-monosomic addition and one monosomic addition were found in the 2n=44 individuals.One double-monosomicsubstitution and one monosomic substitution were obtained.We also proved that it is relatively easier to obtain addition lines in the selfing population than to obtain double-monosomic additions.
, http://www.100md.com
Key words:eremopyrum;wheat;addition;substitution;genomic in situ hybridization(GISH)
旱麦草属[Eremopyrum (L.) Jaub.et Spach]属于禾本科(Gramineae)、小麦族(Triticeae)中的1个1年生属,小麦的野生近缘种,具有生育期短、特早熟、抗旱、耐盐、分蘖力强、抗白粉病等优良性状。所以,开发利用旱麦草属的基因资源无疑对丰富和改善小麦遗传基础具有一定的意义。1993年,刘建文等[1]在国际上首次以普通小麦(Triticum aestivum L.; 2n=6x=42,AABBDD)为母本,以东方旱麦草[Eremopyrum orientale (L.) Jaub.et Spach; 2n=4x=28]为父本,成功地获得了属间远缘杂种F1,并通过幼胚培养,获得了杂种F1植株。同时也对旱麦草属的抗白粉病和耐盐性进行了鉴定。在此基础上我们对普通小麦与东方旱麦草属间杂种的回交2代BC2F1继续进行回交和自交,并对已获得的BC2F3代材料进行了根尖染色体的细胞学及基因组DNA原位杂交检测。
, 百拇医药
1 材料和方法
1.1 材料
小麦-东方旱麦草BC2F3代杂种和东方旱麦草(Er.orientale)种子分别由本实验室选育和保存。
1.2 方法
1.2.1 根尖染色体计数鉴定 种子于常温下浸泡8~10h,25℃暗培养1~2天,截取生长旺盛的根尖在0~4℃冰水中预处理24h,卡诺氏固定液固定24h以上,1%醋酸洋红染色1h,45%醋酸压片,染色体计数,揭片后气干。
1.2.2 染色体制片变性 气干的染色体制片在70℃的70%的甲酰胺、2×SSC溶液中处理2min变性,然后分别在预冷的-20℃的70%、95%、100%的酒精中各脱水5min。
, http://www.100md.com
1.2.3 总基因组DNA的提取和标记 东方旱麦草和中国春的基因组DNA提取用CTAB方法[2],东方旱麦草DNA用biotin-11-dUTP进行缺刻平移法标记[3]。
1.2.4 杂交液配制(8×10μl/片)和杂交液变性 100%甲酰胺40μl;20×SSC(3mol/LNaCl,0.3mol/L柠檬酸钠)8μl;鲑鱼精DNA(1μg/μl)8μl;50%(W/V)硫酸葡聚糖16μl;10% SDS(W/V)0.8μl;探针DNA(1μg/50μl)5μl;封阻DNA(中国春DNA)(0.5μg/μl)10μl;以上杂交液在100℃水浴中保持5min后迅速转入冰浴乙醇中变性处理20min。
1.2.5 杂交 变性处理后的每张染色体制片加10μl变性后的杂交液,加18×18mm盖玻片,放37℃温盒中杂交过夜。
1.2.6 杂交后的漂洗 杂交完毕后,制片标本首先在2×SSC中脱去盖玻片,然后分别在37℃的2×SSC中10min,40℃的2×SSC中5min,25℃的1×PBS中5min。
, http://www.100md.com
1.2.7 荧光信号的检测 每片加0.5%的鼠抗生物素蛋白(第1抗体)50μl,37℃温育1h后,然后分别在37℃ 1×PBS中5min,25℃ 1×PBS中5min。继之,加0.5%连接有荧光素FITC的羊抗鼠抗体(第2抗体)50μl,37℃温育1h后,洗脱同上。最后加7.5μl 1×PI(内含抗褪色剂),封片,观察,照相。
2 结果
2.1 细胞学检测结果
对BC2F3代不同株系抽取的96粒种子的染色体数目进行了细胞学检测,共检测出15粒染色体数目为43的种子,8粒染色体数目为44的种子。其中,由染色体数为43的BC2F2代植株自交产生的BC2F3 46粒种子中,共检测出5粒染色体数为43的类型,占总数的10.87%,2粒染色体数为44,占总数的4.35%。而由染色体数为44的BC2F1代植株经过连续自交两代后产生的BC2F3 50粒种子中,染色体数为43的有10粒,占20%,染色体数为44的有6粒,占总数的12%。其余植株均恢复为小麦正常染色体数2n=42(表1)。
, http://www.100md.com
表1 BC2F3代植株染色体数目
Table1 Chromosome numbers of BC2F3 plants 材料
Materials
不同染色体数目所占频率
No.of chromosomes and frequency
植株总数
No.of plants
42
43
44
, 百拇医药
(BC2F2染色体数为43)
(The number of BC2F2
chromosome is 43)
BC2F3
39(84.78)
5(10.87)
2(4.35)
46
(BC2F2染色体数为44)
(The number of BC2F2
, 百拇医药
chromosome is 44)
BC2F3
34(68.00)
10(20.00)
6(12.00)
50
总数
Total
73(76.04)
15(15.63)
8(8.33)
, http://www.100md.com 96
2.2 基因组DNA原位杂交检测结果(表2)表2 基因组原位杂交检测结果
Table2 Testing results of genotic in situ hybridization 染色体数
No.of
chromosomes
2n=小麦染色体数+
外源染色体数*
2n=No.of wheat chromosome+
alien chromosome*
, 百拇医药
类型
Type
植株数
No.of plants
43
2n=42+1*
单体附加
Monosomic addition
15
44
2n=43+1*
三体、单体附加
, 百拇医药
Wheat trisomic monosomic addition
1
2n=42+2*
二体附加
Disomic addition
5
2n=42+1*+1*
双单体附加
Double-monosomic addition
1
2n=41+3*
, 百拇医药
附加3条外源染色体的单体
Monosomic trible-monosomic addition
1
42
2n=41+1*
单体代换
Monosomic substitution
4
2n=40+1*+1*
双单体代换
Double-monosomic substitution
, 百拇医药
2
*:信号染色体。Alien chromosome
通过基因组DNA原位杂交技术,在染色体数为43的BC2F2植株经自交产生的后代中,检测出1个单体附加(2n=43)(图版I-1),1个二体附加(2n=44)(图版I-2)和1个小麦三体单体附加(2n=44)(图版I-3)。在染色体为44的BC2F1代植株连续自交两代后产生的后代中,检测出1个单体代换(2n=42)(图版I-4),1个双单体代换(2n=42)(图版I-5),2个单体附加(2n=43)(图版I-6,7),1个二体附加(2n=44)(图版I-8),1个附加3条东方旱麦草染色体小麦单体(2n=44)(图版I-9)。另外,在染色体为43的BC2F2植株自交后代中未检测出双单体附加个体,在染色体为44的BC2F1代植株连续自交两代产生的2n=44的5个个体中,检测出1个双单体附加。
, 百拇医药
3 讨论
3.1 小麦-东方旱麦草杂种的配子多样性
通过基因组原位杂交检测出了多种染色体组成方式:染色体数为42的单体代换、双单体代换;染色体数为43的单体附加;染色体数为44的二体附加、双单体附加、附加1条外源染色体的小麦三体和附加3条外源染色体的小麦单体等类型。分析各种不同结果表明,小麦-东方旱麦草的杂种后代减数分裂时形成的配子具有多样性,即小麦染色体在配子中出现了多于或少于正常n=21的情况,产生非正常小麦染色体数目配子的原因很可能是由于外源遗传物质的存在影响了小麦同源染色体的正常配对和分离。我们在BC2F1代花粉母细胞的减数分裂过程中曾观察到了许多落后单价体,并认为,细胞中的单价体数高于含有的外源染色体数,很可能是小麦个别未配对染色体,在减数分裂时,小麦染色体很可能会发生不均衡分离或丢失现象。裴广铮[4]、胡适全[5]分别在小麦×簇毛麦、小麦×苏联球茎大麦杂种后代整倍体株系中观察的结果也表明,外源染色体或其片段的存在影响了小麦染色体的正常配对。东方旱麦草染色体对小麦同源染色体配对频率的影响,有待于进一步研究和探讨。
, 百拇医药
3.2 产生异附加系和代换系的方法讨论
产生异附加系的方法有多种,常规法是用双倍体或部分双倍体回交[6],桥梁亲本法[7,8],通过双单体或多重单体自交法[9,10]可以提高选择频率,胡含1)、翁跃进[11]通过花药培养获得了更高频率的二体附加系。本研究所选得的单体附加和双体附加均是在回交、自交过程中自发形成的。通过细胞学检查,由2n=43的BC2F2代植株自交获得的BC2F3 46粒种子有5粒单体附加,占10.87%,2粒染色体数为44,占4.35%,而由2n=44的BC2F1代植株连续自交2代获得的BC2F3 50粒种子中,有10粒染色体数为43的单体附加,占20.00%;6粒染色体数为44,占12.00%。由于染色体数为44的回交2代(BC2F1)材料未经过自交和人工加倍过程,且我们在其花粉母细胞中观察到的单价体数很高,所以有可能是双单体或多重单体附加植株,由此可见,通过2n=44的植株连续自交选择异附加系的频率明显高于2n=43的植株自交产生附加系的频率。这与Islam[9]、Lukuszewski[10]的观点基本是一致的,但我们的结论的可靠性还有待于进一步加大统计数据去证实。
, 百拇医药
异代换系的常规选育方法是用缺体或单体与附加系杂交再自交产生,张学勇[12]讨论用缺体回交法,陈佩度2)提出用添加二体与单体代换系回交再自交的方法,都可以提高二体代换的频率。本研究所选得的代换材料均是在回交和自交过程中自发产生的,来自于染色体数为44的BC2F1代连续自交两代的材料,这仍然可能是由于杂种后代减数分裂时小麦同源染色体不能正常配对和分离而导致丢失的结果。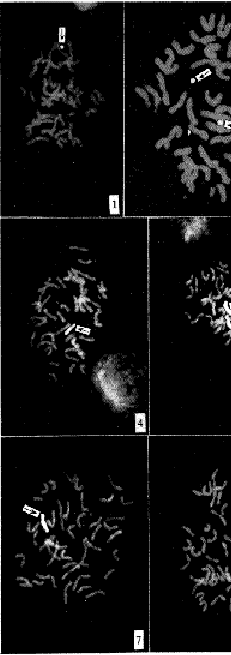

图版说明
1.小麦-东方旱麦草染色体单体附加2n=43;
2.小麦-东方旱麦草染色体二体附加2n=44;
, 百拇医药
3.附加1条东方旱麦草染色体的小麦三体2n=44;
4.小麦-东方旱麦草染色体单体代换2n=42;
5.小麦-东方旱麦草染色体双单体代换2n=42;
6.小麦-东方旱麦草染色体单体附加2n=43;
7.小麦-东方旱麦草染色体双单体代换2n=43;
8.小麦-东方旱麦草染色体二体附加2n=44;
9.附加3条东方旱麦草染色体的小麦单体2n=44以上
箭头所示均为东方旱麦草染色体
Explanation of Plate
, 百拇医药
1.T.aestivum-Er.orientale monosomic addition 2n=43;
2.T.aestivum-Er.orientale disomic addition 2n=44;
3.Wheat trisomic added one orientale chromosome 2n=44;
4.T.aestivum-Er.orientale monosomic substitution 2n=42;
5.T.aestivum-Er.orientale double-monosomic substitution 2n=42;
6.T.aestivum-Er.orientale monosomic addition 2n=43;
, http://www.100md.com
7.T.aestivum-Er.orientale monosomic addition 2n=43;
8.T.aestivum-Er.orientale disomic addition 2n=44;
9.Wheat monosomic added three orientale chromosomes
2n=44.All the arrows shows Er.orientale chromosomes
参考文献
[1]Liu J W et al.Induction of immature embryo callus,establishment of somaclones and regeneration of plant from Triticum aestivum×Eremopyrum orientale hybird.Proc.8th Int.Wheat Gen.Syn.,China Agr.Sci.Press,1995,Vol(1):217~222.
, 百拇医药
[2]Saghai-Maroof M A et al.Ribosomal DNA Length polymorphism in barley:Mendelian inheritance chromosomal location and population dynamics.Proc.Natl.Acad.Sci.USA,1984,81:8014~8018.
[3]Friebe B et al.Radiation-induced nonhomoeologous wheat-Agropyron intermedium chromosomal translocations conferring resistance to leaf rust.Thero.Appl.Genet.,1993,86:141~149.
[4]裴广铮等.小麦与簇毛麦杂种后代抗白粉病株系的细胞遗传学分析.南京农业大学学报,1986,(1):1~9.
, 百拇医药
[5]胡适全等.普通小麦(6x)×苏联球茎大麦(4x)属间整倍体杂种后代的细胞遗传学研究. 遗传学报,1990,18(1):67~72.
[6]Riley R et al.Introduction of yellow rust resistance of Aegilops comosa into wheat by genetically induced homoeologous recombination.Nature,1968,5126(217):383~384.
[7]刘大钧等.将簇毛麦种质转移给小麦的研究.遗传学报,1983,10(2):103~113.
[8]Hyde Beal B.Addition of individual Haynaldia villosa chromosomes to hexaploid wheat.American Journal of Botany,1953.40(3):174~182.
, http://www.100md.com
[9]Islam A K M R et al.Production and characterization of wheat-barley addition lines.Proc.5th Int.Wheat Genet.Symp.,New Delhi.1978,365~371.
[10]Lukaszewskei A.J.Acomparison of Several approaches in the development of disomic alien addition lines of wheat.Proc.7th Int.Wheat Genet.Symp.,Cambridge,England.1988,363~367.
[11]翁跃进等.普通小麦-顶芒山羊草异源附加系的创建和鉴定.Ⅰ.小麦花药培养对创建普通小麦-顶芒山羊草异源附加系的作用.作物学报,1995,21(1):39~44.
[12]张学勇等.缺体回交法选育普通小麦异代换系方法的研究.遗传学报,1989,16(6):420~429., 百拇医药
单位:张桂芳 刘建文 黄远樟 丁敏(北京师范大学生命科学学院,北京 100875);唐顺学 贾旭(中国科学院遗传研究所,北京 100101)
关键词:旱麦草;普通小麦;附加系;代换系;基因组原位杂交(GISH)
遗传学报000109
摘要:旱麦草属(Eremopyrum)是用于小麦品种改良的又一潜在的植物资源。为了筛选小麦-旱麦草异附加系、异代换系,对普通小麦品种Fukuho×东方旱麦草属间杂种的BC2F3代材料的96粒种子进行了染色体数目的检测,共检出15粒2n=43的种子,8粒2n=44的种子,进一步对以上材料进行的基因组DNA原位杂交,共鉴定出3个单体附加系,2个二体附加系,1个双单体附加,1个小麦三体单体附加,1个附加3条东方旱麦草染色体的小麦单体,在染色体数为42的个体中,检测出1个单体代换,1个双单体代换。根据BC2F3代自交品系来源的不同,初步认为由双单体附加自交比单体附加自交选择异附加系的效率高。
, http://www.100md.com
中图分类号:Q343 文献标识码:A
文章编号:0379-4172(2000)01-0050-0055
Isolation and Verification of Triticum aestivum-Eremopyrum orientale Addition Lines and Substitution Lines by GISH
ZHANG Gui-Fang,LIU Jian-Wen,HUANG Yuan-Zhang,DING Min
(College of Life Sciences,Beijing Normal University,Beijing 100875,China)
TANG Shun-Xue,JIA Xu
, http://www.100md.com
(Institute of Genetics,Chinese Academy of Sciences,Beijing 100101,China)
Abstract:Eremopyrum is a potential genus in wheat improvement.To breed T.aestivum-Er.orientale additional and substitutional lines,chromosome number of 96 BC2F3 individuals were accounted.Fifteen plants with 2n=43 and eight plants with 2n=44 were checked out.GISH results of the individuals with 43 chromosomes showed that they blonged to three different monosomic additions.Two disomic additions,one double-monosomic addition,one trible-monosomic addition and one monosomic addition were found in the 2n=44 individuals.One double-monosomicsubstitution and one monosomic substitution were obtained.We also proved that it is relatively easier to obtain addition lines in the selfing population than to obtain double-monosomic additions.
, http://www.100md.com
Key words:eremopyrum;wheat;addition;substitution;genomic in situ hybridization(GISH)
旱麦草属[Eremopyrum (L.) Jaub.et Spach]属于禾本科(Gramineae)、小麦族(Triticeae)中的1个1年生属,小麦的野生近缘种,具有生育期短、特早熟、抗旱、耐盐、分蘖力强、抗白粉病等优良性状。所以,开发利用旱麦草属的基因资源无疑对丰富和改善小麦遗传基础具有一定的意义。1993年,刘建文等[1]在国际上首次以普通小麦(Triticum aestivum L.; 2n=6x=42,AABBDD)为母本,以东方旱麦草[Eremopyrum orientale (L.) Jaub.et Spach; 2n=4x=28]为父本,成功地获得了属间远缘杂种F1,并通过幼胚培养,获得了杂种F1植株。同时也对旱麦草属的抗白粉病和耐盐性进行了鉴定。在此基础上我们对普通小麦与东方旱麦草属间杂种的回交2代BC2F1继续进行回交和自交,并对已获得的BC2F3代材料进行了根尖染色体的细胞学及基因组DNA原位杂交检测。
, 百拇医药
1 材料和方法
1.1 材料
小麦-东方旱麦草BC2F3代杂种和东方旱麦草(Er.orientale)种子分别由本实验室选育和保存。
1.2 方法
1.2.1 根尖染色体计数鉴定 种子于常温下浸泡8~10h,25℃暗培养1~2天,截取生长旺盛的根尖在0~4℃冰水中预处理24h,卡诺氏固定液固定24h以上,1%醋酸洋红染色1h,45%醋酸压片,染色体计数,揭片后气干。
1.2.2 染色体制片变性 气干的染色体制片在70℃的70%的甲酰胺、2×SSC溶液中处理2min变性,然后分别在预冷的-20℃的70%、95%、100%的酒精中各脱水5min。
, http://www.100md.com
1.2.3 总基因组DNA的提取和标记 东方旱麦草和中国春的基因组DNA提取用CTAB方法[2],东方旱麦草DNA用biotin-11-dUTP进行缺刻平移法标记[3]。
1.2.4 杂交液配制(8×10μl/片)和杂交液变性 100%甲酰胺40μl;20×SSC(3mol/LNaCl,0.3mol/L柠檬酸钠)8μl;鲑鱼精DNA(1μg/μl)8μl;50%(W/V)硫酸葡聚糖16μl;10% SDS(W/V)0.8μl;探针DNA(1μg/50μl)5μl;封阻DNA(中国春DNA)(0.5μg/μl)10μl;以上杂交液在100℃水浴中保持5min后迅速转入冰浴乙醇中变性处理20min。
1.2.5 杂交 变性处理后的每张染色体制片加10μl变性后的杂交液,加18×18mm盖玻片,放37℃温盒中杂交过夜。
1.2.6 杂交后的漂洗 杂交完毕后,制片标本首先在2×SSC中脱去盖玻片,然后分别在37℃的2×SSC中10min,40℃的2×SSC中5min,25℃的1×PBS中5min。
, http://www.100md.com
1.2.7 荧光信号的检测 每片加0.5%的鼠抗生物素蛋白(第1抗体)50μl,37℃温育1h后,然后分别在37℃ 1×PBS中5min,25℃ 1×PBS中5min。继之,加0.5%连接有荧光素FITC的羊抗鼠抗体(第2抗体)50μl,37℃温育1h后,洗脱同上。最后加7.5μl 1×PI(内含抗褪色剂),封片,观察,照相。
2 结果
2.1 细胞学检测结果
对BC2F3代不同株系抽取的96粒种子的染色体数目进行了细胞学检测,共检测出15粒染色体数目为43的种子,8粒染色体数目为44的种子。其中,由染色体数为43的BC2F2代植株自交产生的BC2F3 46粒种子中,共检测出5粒染色体数为43的类型,占总数的10.87%,2粒染色体数为44,占总数的4.35%。而由染色体数为44的BC2F1代植株经过连续自交两代后产生的BC2F3 50粒种子中,染色体数为43的有10粒,占20%,染色体数为44的有6粒,占总数的12%。其余植株均恢复为小麦正常染色体数2n=42(表1)。
, http://www.100md.com
表1 BC2F3代植株染色体数目
Table1 Chromosome numbers of BC2F3 plants 材料
Materials
不同染色体数目所占频率
No.of chromosomes and frequency
植株总数
No.of plants
42
43
44
, 百拇医药
(BC2F2染色体数为43)
(The number of BC2F2
chromosome is 43)
BC2F3
39(84.78)
5(10.87)
2(4.35)
46
(BC2F2染色体数为44)
(The number of BC2F2
, 百拇医药
chromosome is 44)
BC2F3
34(68.00)
10(20.00)
6(12.00)
50
总数
Total
73(76.04)
15(15.63)
8(8.33)
, http://www.100md.com 96
2.2 基因组DNA原位杂交检测结果(表2)表2 基因组原位杂交检测结果
Table2 Testing results of genotic in situ hybridization 染色体数
No.of
chromosomes
2n=小麦染色体数+
外源染色体数*
2n=No.of wheat chromosome+
alien chromosome*
, 百拇医药
类型
Type
植株数
No.of plants
43
2n=42+1*
单体附加
Monosomic addition
15
44
2n=43+1*
三体、单体附加
, 百拇医药
Wheat trisomic monosomic addition
1
2n=42+2*
二体附加
Disomic addition
5
2n=42+1*+1*
双单体附加
Double-monosomic addition
1
2n=41+3*
, 百拇医药
附加3条外源染色体的单体
Monosomic trible-monosomic addition
1
42
2n=41+1*
单体代换
Monosomic substitution
4
2n=40+1*+1*
双单体代换
Double-monosomic substitution
, 百拇医药
2
*:信号染色体。Alien chromosome
通过基因组DNA原位杂交技术,在染色体数为43的BC2F2植株经自交产生的后代中,检测出1个单体附加(2n=43)(图版I-1),1个二体附加(2n=44)(图版I-2)和1个小麦三体单体附加(2n=44)(图版I-3)。在染色体为44的BC2F1代植株连续自交两代后产生的后代中,检测出1个单体代换(2n=42)(图版I-4),1个双单体代换(2n=42)(图版I-5),2个单体附加(2n=43)(图版I-6,7),1个二体附加(2n=44)(图版I-8),1个附加3条东方旱麦草染色体小麦单体(2n=44)(图版I-9)。另外,在染色体为43的BC2F2植株自交后代中未检测出双单体附加个体,在染色体为44的BC2F1代植株连续自交两代产生的2n=44的5个个体中,检测出1个双单体附加。
, 百拇医药
3 讨论
3.1 小麦-东方旱麦草杂种的配子多样性
通过基因组原位杂交检测出了多种染色体组成方式:染色体数为42的单体代换、双单体代换;染色体数为43的单体附加;染色体数为44的二体附加、双单体附加、附加1条外源染色体的小麦三体和附加3条外源染色体的小麦单体等类型。分析各种不同结果表明,小麦-东方旱麦草的杂种后代减数分裂时形成的配子具有多样性,即小麦染色体在配子中出现了多于或少于正常n=21的情况,产生非正常小麦染色体数目配子的原因很可能是由于外源遗传物质的存在影响了小麦同源染色体的正常配对和分离。我们在BC2F1代花粉母细胞的减数分裂过程中曾观察到了许多落后单价体,并认为,细胞中的单价体数高于含有的外源染色体数,很可能是小麦个别未配对染色体,在减数分裂时,小麦染色体很可能会发生不均衡分离或丢失现象。裴广铮[4]、胡适全[5]分别在小麦×簇毛麦、小麦×苏联球茎大麦杂种后代整倍体株系中观察的结果也表明,外源染色体或其片段的存在影响了小麦染色体的正常配对。东方旱麦草染色体对小麦同源染色体配对频率的影响,有待于进一步研究和探讨。
, 百拇医药
3.2 产生异附加系和代换系的方法讨论
产生异附加系的方法有多种,常规法是用双倍体或部分双倍体回交[6],桥梁亲本法[7,8],通过双单体或多重单体自交法[9,10]可以提高选择频率,胡含1)、翁跃进[11]通过花药培养获得了更高频率的二体附加系。本研究所选得的单体附加和双体附加均是在回交、自交过程中自发形成的。通过细胞学检查,由2n=43的BC2F2代植株自交获得的BC2F3 46粒种子有5粒单体附加,占10.87%,2粒染色体数为44,占4.35%,而由2n=44的BC2F1代植株连续自交2代获得的BC2F3 50粒种子中,有10粒染色体数为43的单体附加,占20.00%;6粒染色体数为44,占12.00%。由于染色体数为44的回交2代(BC2F1)材料未经过自交和人工加倍过程,且我们在其花粉母细胞中观察到的单价体数很高,所以有可能是双单体或多重单体附加植株,由此可见,通过2n=44的植株连续自交选择异附加系的频率明显高于2n=43的植株自交产生附加系的频率。这与Islam[9]、Lukuszewski[10]的观点基本是一致的,但我们的结论的可靠性还有待于进一步加大统计数据去证实。
, 百拇医药
异代换系的常规选育方法是用缺体或单体与附加系杂交再自交产生,张学勇[12]讨论用缺体回交法,陈佩度2)提出用添加二体与单体代换系回交再自交的方法,都可以提高二体代换的频率。本研究所选得的代换材料均是在回交和自交过程中自发产生的,来自于染色体数为44的BC2F1代连续自交两代的材料,这仍然可能是由于杂种后代减数分裂时小麦同源染色体不能正常配对和分离而导致丢失的结果。
图版说明
1.小麦-东方旱麦草染色体单体附加2n=43;
2.小麦-东方旱麦草染色体二体附加2n=44;
, 百拇医药
3.附加1条东方旱麦草染色体的小麦三体2n=44;
4.小麦-东方旱麦草染色体单体代换2n=42;
5.小麦-东方旱麦草染色体双单体代换2n=42;
6.小麦-东方旱麦草染色体单体附加2n=43;
7.小麦-东方旱麦草染色体双单体代换2n=43;
8.小麦-东方旱麦草染色体二体附加2n=44;
9.附加3条东方旱麦草染色体的小麦单体2n=44以上
箭头所示均为东方旱麦草染色体
Explanation of Plate
, 百拇医药
1.T.aestivum-Er.orientale monosomic addition 2n=43;
2.T.aestivum-Er.orientale disomic addition 2n=44;
3.Wheat trisomic added one orientale chromosome 2n=44;
4.T.aestivum-Er.orientale monosomic substitution 2n=42;
5.T.aestivum-Er.orientale double-monosomic substitution 2n=42;
6.T.aestivum-Er.orientale monosomic addition 2n=43;
, http://www.100md.com
7.T.aestivum-Er.orientale monosomic addition 2n=43;
8.T.aestivum-Er.orientale disomic addition 2n=44;
9.Wheat monosomic added three orientale chromosomes
2n=44.All the arrows shows Er.orientale chromosomes
参考文献
[1]Liu J W et al.Induction of immature embryo callus,establishment of somaclones and regeneration of plant from Triticum aestivum×Eremopyrum orientale hybird.Proc.8th Int.Wheat Gen.Syn.,China Agr.Sci.Press,1995,Vol(1):217~222.
, 百拇医药
[2]Saghai-Maroof M A et al.Ribosomal DNA Length polymorphism in barley:Mendelian inheritance chromosomal location and population dynamics.Proc.Natl.Acad.Sci.USA,1984,81:8014~8018.
[3]Friebe B et al.Radiation-induced nonhomoeologous wheat-Agropyron intermedium chromosomal translocations conferring resistance to leaf rust.Thero.Appl.Genet.,1993,86:141~149.
[4]裴广铮等.小麦与簇毛麦杂种后代抗白粉病株系的细胞遗传学分析.南京农业大学学报,1986,(1):1~9.
, 百拇医药
[5]胡适全等.普通小麦(6x)×苏联球茎大麦(4x)属间整倍体杂种后代的细胞遗传学研究. 遗传学报,1990,18(1):67~72.
[6]Riley R et al.Introduction of yellow rust resistance of Aegilops comosa into wheat by genetically induced homoeologous recombination.Nature,1968,5126(217):383~384.
[7]刘大钧等.将簇毛麦种质转移给小麦的研究.遗传学报,1983,10(2):103~113.
[8]Hyde Beal B.Addition of individual Haynaldia villosa chromosomes to hexaploid wheat.American Journal of Botany,1953.40(3):174~182.
, http://www.100md.com
[9]Islam A K M R et al.Production and characterization of wheat-barley addition lines.Proc.5th Int.Wheat Genet.Symp.,New Delhi.1978,365~371.
[10]Lukaszewskei A.J.Acomparison of Several approaches in the development of disomic alien addition lines of wheat.Proc.7th Int.Wheat Genet.Symp.,Cambridge,England.1988,363~367.
[11]翁跃进等.普通小麦-顶芒山羊草异源附加系的创建和鉴定.Ⅰ.小麦花药培养对创建普通小麦-顶芒山羊草异源附加系的作用.作物学报,1995,21(1):39~44.
[12]张学勇等.缺体回交法选育普通小麦异代换系方法的研究.遗传学报,1989,16(6):420~429., 百拇医药