TCR-BCRʶ��ԭ�����ڷ�����̬ˮƽ�ıȽ�
���ߣ������ ��ѧ�� �� �� �� �� ������ ������w'(y/1, ��Ĵҽҩ
��λ������ҽ�ƴ�ѧ����ѧ�о��� �人 430071w'(y/1, ��Ĵҽҩ
�ؼ��ʣ�w'(y/1, ��Ĵҽҩ
�� �� ѧ �� ־990301COMPARISON OF TCR-BCR ANTIGEN RECOGNITION MECHANISMw'(y/1, ��Ĵҽҩ
AT THE MOLECULAR ECOLOGICAL LEVELw'(y/1, ��Ĵҽҩ
��������ѧ��������ķ�չ�У������е��˱�����������ʵ���ˮƽ�ͳ���ˮƽ��������ı��ʣ�Ҳ�����������ڵĻ����������ṹ���ܣ�����ˮƽ�Թ⾵��ϸ��ѧ���漼��Ϊ���ڳ���ˮƽ�Ե羵�ͷ�������ѧ����Ϊ������ѧ��������ѧ�е�һ����Ҫ����һ��ѧ�Ƶķ�չ���������Ź���ĸı�Ͷ���������ʶ������ѧ�Բ����⡣�����Ƕ�����ѧ�ļ�ʶ�������ɼ�һ������ѧ����ϱ仯��չ���������ҹ���1964������˷�������ѧ�ĸ���[1]���ڵ�ʱ��˵�ǶԴ�ͳ����ѧ��һ�������Ըı䡣16���������������ѧ�����ԡ�����ѧ�ľ����ԺͷǾ����ԡ�Ϊ�⣬�Դ�ͳ������ѧ�������ߺͷ��������ߣ������������߸�����ᷨ�ϲ������������[2]���Ⱥ�õ���������һ������һ��Ȩ������ѧ��л���ĺ��������ȵ�֧�ֺ����͡��������ܵ�������̬ѧ��չ�������������ڹ����������������̬ѧ������[3]֮��������ȻҪ����̬ѧ�������뵽��������ѧ��������1993�������ˡ������صķ�����̬ѧ�������ķ���������ѧ��־������������ѧ��ժ(CA)ժ��[4]���ڿ����ǹ��������ġ�������̬ѧ����3ƪ�У���9��(16��24��)���������߷�����̬ѧ���й�����[5]������˵һ��ո�µ�����ѧ��֧ѧ��(������̬����ѧ)�Ѿ����������������Dz��϶����ķ�չ�����ʶ������̽�֡������Զ������û������кܶ��˽��TCR(Tϸ������)��BCR(Bϸ������)�Ĵ�ͳ������з�����̬ѧ�ķ����Ƚϣ����ѷ������߷�Ӧ�������غ�Эͬ����֮������ӹ�ϵ���ƺ����ģ�������ˣ����������������ӹ�ϵ����νģ���Խ������ۣ�����������ģ�������֮��w'(y/1, ��Ĵҽҩ
1 ʲô��T1Bϸ��ʶ����ԭ����������w'(y/1, ��Ĵҽҩ
T��Bϸ��ʶ��ԭ��������ӵ�Ȼ��TCR��BCR���������ڸ���ϸ���ı��档�����ǵ��������õ����֣������������������ͱ�����ӵ�Эͬ������CD����ϵͳ(Cluster desigation)�ȡ��������ֵĿ�ԭʶ�����������ܵ�MHC/��ԭ�ĸ�������Ϊ��������֮�⣬��Ҫ�ܵ�CD3��CD4��CD8�ȱ�����ӵ�Ӱ�죬CD3��TCR���ӵ���ɲ��֣�CD4��CD8�ֱ����Tϸ���Ĺ�������������ߵ�����(T����ϸ����Th)������ɱ�����ߵ�����(Tɱ��ϸ����Tk����ϸ����ϸ����TC)������һ����ΪTϸ����ϸ�����ߵ����壬������������ʶ��ԭ�ĵķ�����TCR���ⱻ��Ϊ����̼������ڷ��������߹���ʱ�����������CD4��CD8���ӵĴ̼�����ⱻ��ΪЭͬ�̼��������ǹ��ϵ�ϸ�����ߵķ��ӻ��ƣ�Ҳ��������ָ���������ӣ���ȻTCR������CD4��CD8�Ǵӡ��ӷ�����̬ѧ�ĸ�������������TCR��CD4��CD8��һ��(TCR-CD4��TCR-CD8)������Է��ӵĹ��ܿ���������Ϊ���ӷ����ǻ�Ϊ���������ģ�CD4��CD8����������Եı��֣�����������TCR�Ĵ��ڣ����������������Tϸ�����߷�Ӧ�е����þͲ����ܱ��ֳ�����TCRʶ�������Կ�ԭ�������ԣ�Ҳ����������CD4��CD8��Ϊ��������������Э��������TCR��������ϸ�����߹���Ҳ�����ܱ��ֳ�����������Dz���ð������������Է��ӵ���̬�Ե��¸���Ա���������̬ѧͬ��������ѧ��������֮�㣬�����ڷ�������ѧֻ��������Է��ӵ������ԣ�������̬ѧ��������Է��ӵ���̬�ԡ���̬�Ծ���ָ����ͬ������ͳһ���ں��ˮƽ������ĸ�����Ⱥ�壬�Ի����������������Ǿ��Եģ�����(ϸ��)ˮƽ�����ֹ�����ϸ���������书�ܻ��Ȼ���ض��Ļ����������ڳ���(����)ˮƽ�����ַ����������������Է��ӣ�Ҳ��Ȼ�����ض��ķ�����̬���������ǶԷ�����̬ѧ���������ᷨ���ɼ�Ϊ��������̬ѧ���о�������Է�����̬�ԵĿ�ѧ�����߷�����̬ѧ���о�������������Է�����̬�ԵĿ�ѧ����ϣ���������ۣ�����ӭ������
2 BCR�ṹ�빦�ܵ���̬��a&7n, ��Ĵҽҩ
Bϸ������Ȼ��ԭ��ʶ��ͷ�Ӧ�������ڱ������������ߵ�BCR��BCR�ܿ�ԭ�̼�������İ����ٲ��¼����ɵ���ϸ���Ļ�ͷֻ���ֱ������(anergy)����������ʼ����BCR�̼����Ե����Ұ�����øϵͳ���˼�ø������PLCr-1��2(��֬øCr-1��2)�����ữ�ͻ��ͬʱras����ͬShc��Grb2��mSOS����ɻ������(activation complex)���Ӷ�ʹRaf���ャĤ����˿/�հ�����ø��ٲ�������ϸ����ֳ�� ����Ӧ������߷�Ӧ��������Ҫ��a&7n, ��Ĵҽҩ
��ͳ�������������Һ���ߣ�ָ���ǿ�����ӣ����屾����λ��Bϸ��Ĥ�����������(Ig��immunoglobulin)���������Ե�һ���Ӵ��ڣ�ʵ����һ����˫����������ǵ���˫�壬��Ig����Ig�·ֱ���mb-1��B29������룬���߶��ǿ�Ĥ�ṹ������Ĥ��������Ĥ���Ͱ��������������ڵ�������(AA)����Ig��Ϊ61��Ig��Ϊ48AA��Ig����Ig�º��г�Ϊ��ԭʶ������(antigen recognition activation motif�� ARAM)��ƿ�ԭʶ��ͬ����-1(antigen recognition homology 1��ARH1)�������������ѷ����������ͼ1��ʾ�� a&7n, ��Ĵҽҩ
a&7n, ��Ĵҽҩ
ͼ1 Ig����Ig��AA���н�Ͽ�ԭ���ӱȽ�a&7n, ��Ĵҽҩ
����ʾAA�л���������ʾ������������Ҳ��֮�ı�a&7n, ��Ĵҽҩ
-��ʾ�����Բл�a&7n, ��Ĵҽҩ
ͼ1ʾBCR����������Ig����Ig�µ�AA���У�������ͬ��ӦЧӦ�������ϵ��������У�������������ARAM��ARH-1��Ig����Ig�¶�����ARH-1����ͬЧӦ�������ϵ������Ը��Բ�ͬ��Ig����DCSM���Ķ�ͬSrc���弤øLyn��Fyn��Blk��P38KDa��ϣ���Ig�º�QTAT���Ķ������������ֵ���40��42KDa��ͼ1����ʾ��ͨ��ת��������֤��Ig����DCSM��Ig�µ�QTAT�û�����Ig����ͬЧӦ���ӽ�������Ա�ΪIg���͵������ԣ���֮��Ȼ����Ig�����ԣ��б������[6]��YEGLN��YEDI�������ữʱ�������Ӷ�Fyn�Ľ����3���������߶������ữʱ������������20���������Ұ������ữ��ЭͬЧӦ(synergy effect)ָʾ��ЧӦ���ӽ�ϵĺ�����(co-operativity)��
Tseng��[7] �������ͷ���ģʽ�ر�������Ig�����ӹ��͵ľ�ֹ״̬��״̬����ʾ�ķ������������̬�Ա仯������ƣ�Tseng�������ģʽͼ��������Ϊͼ2�����Ա��ֳ�������̬�Եĺ��⣬��һģʽͼ��������Ϊͼ3������BCR������ʱ��Ϊ���ӵķ�����̬��������ϵ�� mq{&$, ��Ĵҽҩ
mq{&$, ��Ĵҽҩ
ͼ2 BCR Ig���Ĺ��ͱ仯mq{&$, ��Ĵҽҩ
ͼ2ʾmIg BCR�İ���������˫��Ig����Ig�µĹ���ϵ����ʾ��ֹ̬����øFyn��N-�����ӽ��棬��ͬIg����DCSM��ϣ���SH2����ĩ��ø�������BCR�ܿ�ԭ�̼����Ig���������Ըı䣬��������CD45�������ã���Ϊ�Ұ�������ø(tyrosine phosphatase)���������������ã�һ�Ǵ�Fyn���۵������Ǵٽ�Fyn�ļ�ø���ԣ����ȶ���״̬��ͬʱʹIg����YEDI��YEGLNȥ���ữ�����⣬CD45ͨ��Ig��/Ig�����ữ��ȥ���ữ���̣����ܻ��дٽ��źŴ��ݵ����á�mq{&$, ��Ĵҽҩ
��һ���������Ѿ���ļ�ø��Fyn�ȼ���������BCR�������ϣ���Ϊͬ�����μ�ЧӦ����(secondary effectors)��ϵ����������磬Fyn��SH2�������Ig������SH3��������P13-k�����䰱�����ֽ����PLCr(phospholipase Cr)[8]������ͬIg������ϵ��FynͬP13-k��PLCr���ϵ���ø�������磬�������̬ϵ���临��Ҳ���ο�ͼ3�� mq{&$, ��Ĵҽҩ
mq{&$, ��Ĵҽҩ
ͼ3 һ����BCR���帴����ģʽmq{&$, ��Ĵҽҩ
ͼ3ʾBCR�������Ӹ�������γɡ�ͬ�����ܷ�����ϵ��ЧӦ���ӣ�Ҳ��ͬ���������帴������ɰ�������(intracytoplasmic bridges)���źŴ��ݷ���Ӱ�졣����P13-k����ֻͬ����������Fyn��Lynͨ��SH3������ϵ����������SH2 ͬCD19���ӷ�����ϵ��ͼ3��ȷָ����BCR����������3���źŴ���;����������̼��������Ұ��ἤø�������Щ��ļ�ø������ʹIg����Ig�����ữ����ֱ�ӵĻ��ӵ�ʹCD19��CD22Ҳ���ữ������Щ���Ұ����Χ�����ǵ����ж�����ΪЧӦ������ʾ���ʹ������������Ĵμ��������źŴ��ݷ���(secondary and tertiary signal tranductors)�����á�����ʵ�������Bϸ������CD22�ı���Ͳ���ͨ��BCR�����ź�[9]����һ��Է�����̬����ѧ���ԣ��ر������壬��Ϊһ������ȱʧ(��һ������������ȱ��)�Ϳ�ʹ����Ӧ�еĹ��ܣ��������ܡ�
ͼ3�е�CD19��Ϊ�����ף�ͬCD21��ɸ����壬CD21���Dz�������2(CR2)����һ�����廹����TAPA���Ϸ��ӣ�CD19�ڽ���ΪSH2�ṩ������ϵ㣬����ͬ�������ӵ�������ϵ��h^9hcq, ��Ĵҽҩ
CD22Ϊ˫�壬��ΪCD22a��CD22b������������ͬԴ�����ԣ���3��ճ�Է��ӣ����走�ס���ϸ��ճ�Է��Ӻ�Ѫ��ճ�Է���(VCAM-1)��CD22a�鵼Bϸ��ճ���ڵ���ϸ���ͺ�ϸ����(��ϸ������)��CD22b����B-B��B-T֮����ճ�����á�h^9hcq, ��Ĵҽҩ
�����ǹ���BCR������̬�Եļ�Ҫ�����չ��δ�����ؽ����µĽ�չ����Ŀ�Դ���Ҳ��h^9hcq, ��Ĵҽҩ
3 TCR�ṹ�빦�ܵ���̬��h^9hcq, ��Ĵҽҩ
��ͳ��TCRʶ��ԭ�������Ի��ƣ�һֱ���DZȶ�BCR�ܵ��������ӵ����⣬���ﲻ������ϸ�����ۣ�ֻ�轫Koretzky[10]��Ward SG��[11]����Tϸ���������ʾ��ͼ���ۺ�Ϊ���ĵ�ͼ4���ٸ���˵�����ɣ������ǽ�ǿ��ijЩЭͬ���Ӷ�TCR���ܱ����е���Ҫ���壬����������BCR�Ľṹ����ʱ��Ҳ��ǿ������һ�㡣 h^9hcq, ��Ĵҽҩ
h^9hcq, ��Ĵҽҩ
ͼ4 TCR�Ľṹ�빦�ܼ�CD28������ʾ��h^9hcq, ��Ĵҽҩ
ERK��extracellular signal-regulated kinaseh^9hcq, ��Ĵҽҩ
FRAP��FK506-binding protein rapamycin associated proteinh^9hcq, ��Ĵҽҩ
JNK��c-jun N-terminal kinaseh^9hcq, ��Ĵҽҩ
PI3-kinase(PI3K)��phospoinositide 3-kinaseh^9hcq, ��Ĵҽҩ
PKC��protein kinase Ch^9hcq, ��Ĵҽҩ
PLC��phosphalipase Ch^9hcq, ��Ĵҽҩ
PtdIns(3��4��5)P3��phosphotidylinositol(3��4��5)-triphsphate
PTK��protein tryosine kinasdo, ��Ĵҽҩ
Ras��P21rasdo, ��Ĵҽҩ
SM��acidic sphingomeyelinasedo, ��Ĵҽҩ
TCR��T-cell receptordo, ��Ĵҽҩ
TOR��target of rapamycindo, ��Ĵҽҩ
TCRʵ���Ǧ�������ͬCD3 �ܦš��Ħż�˫�ɦ���ɵĸ����壬��δע�⸴���������ɵ���̬�����塣��ȻCD4��CD8����ΪTϸ����ͬ���ܱ��ֵı�Ƿ��ӣ��ܵ���һ���̶ȵ�ע�⣬Ҳ��δ�����ǿ��CD4��CD8��Tϸ���Ĺ��ܱ����ʵ���ž����Ե����ã�����������һ����Ϊ����CD4�����Tϸ���Dz�����ʹ������������õģ���CD�����Tϸ����Ҳ�Dz�����ʹ��ɱ�˰�ϸ�������õģ�����TCR�Ĺ��ܱ��֣�����ϵ��������CD8������CD28(ͼ4)��CD2�ȡ�TCR������Ҫ�źŴ���·���Ǧ�LC/Calcium P21ras�� PI3k�� P21ras��Ras����TCR�Ļ���CD28��P13k��ø���Ӷ�����CD28��TCR֮��ġ���̸��(cross talk)��P13k�����ܼ����ź����ε�Ras��Rac��Pkc��Bcl-XL�������P7056kҲ�����TCR���Ŀ��ͨ�����ӵ��ź�(���ǿɳ�֮Ϊ������̬����)�������ڻ�ʹٽ�IL-2�ı���Ӷ������ͼ�ǿTϸ���������ԡ�do, ��Ĵҽҩ
TCR�Ĺ��ܱ�����������ᵽ��CD28�⣬CD2�����ã�Ҳ���ܶ�֮����Ҫ��Ӱ�죬����Davis�ȶԴ������൱��ϸ�Ľ���[11]��CD2ʵ����һ������[����CD58��CD48��SLAM(T)��2B4(NK)��Ly-9(T2B)]����CEA(���߿�ԭ)����[ ����CD66c(NCA)��CD66b(CGMb)��CD66l(CEA)]ͬ��IgSF(�����׳�����)��ͼʾ����(ͼ5)���������ˡ� do, ��Ĵҽҩ
do, ��Ĵҽҩ
ͼ5 CD2��CEA��ΪIgSF�����������Աʾ��do, ��Ĵҽҩ
������Ĥ��
����glycosylphosphatidylinositol�� GPI��ʾí������l, ��Ĵҽҩ
TCRʶ��APC(antigen presentate cell)������ӳʵݵ�MHC-��ԭ��ʱ��CD2ͬAPC�ϵ�����CD58(LFA-3)�ܰ�ϸ�����ܷ���-3)��ϣ��м�ǿMHC���Ƶ�Tϸ��ʶ��ԭ�ĵ����á�Tϸ��ʶ��ԭ�Ĺؼ�������Tϸ����APC�ĽӴ�������֮���ж������͵ķ������ϣ���������ϸ���ϱ�����ӵ����Ͽ���Ϊ�ǻ�Ϊ����������Ľ�ϣ��м�ǿϸ����ϸ����ճ�����ã�ϸ��ճ�Է�����ϸ���������е���̬�Է���ֻ����ϸ�����߾����Ը����о������⣬�������߷�����̬ѧ�ľ������ݡ�Davis��������CD2������ʱ����ͼ����ʽ������ÿ�ַ��ӵ�ճ�����ã�����Ϊ����ͼ6a��b��c�����ʲο���l, ��Ĵҽҩ
ͼ6a ʾTϸ������ķ������Ƽ��������Сl, ��Ĵҽҩ
ITAM��immunoreceptor tyrosine based activation motifl, ��Ĵҽҩ
GPI����ͼ5l, ��Ĵҽҩ
����Ҫָ�����ǣ�TCRCD3�����庬�����������Ұ�������(ITAM)���ڽ�CD2���ӣ�CD2��TCR��������ݽᾧѧ�͵羵�����о�������ʾ����CD45��CD43ҪС6��8���������ǵı����ܶ��ֱ�TCR��CD2��2��3��(ͼ6a��b)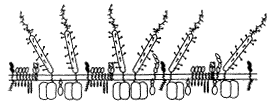 l, ��Ĵҽҩ
l, ��Ĵҽҩ
ͼ6b ʾ��ֹ��Tϸ���ı�����ӣ��ص�������ʾCD2��CD45��CD4��TCRCD3�ȵķֲ���ϵl, ��Ĵҽҩ
CD2ͬABC��������ʱ���ɵ���T��APC��ϸ��Ĥ���ܽӴ�������CD45�ķ��ӽϴ����Ե�TCRͬMHC/��ԭ�ĺ�CD2ͬ������ʱ�����뽫CD45�������Ա��ſ�������������ϸ������Ӵ�����ͼ6c���Կ���CD2/����ͬTCR/MHC��ԭ�ĸ�������ɸ����ӵĸ�����ϵͳ���縴�����������������������CD4���Ӳ�ͬLck��ø����ϵ��ֻ�и�������TCR��ʶ��ԭ�ģ��Ӷ�ʹLck����ITAM���ữ���ã�����һ�����ZAP-70���������ữ����źţ�����TCR���������Եijɹ��������Ӧ�ñ�����IL-2�Ļ�����IL-2�����ϡ������������CD4ȱʧ��MHCδ�п�ԭ�Ľ�ϣ���Ȼ���ᷢ���������ã������������CD4�Ϳ�ԭ�Ĵ��ڣ�����CD2/����IJ��룬Ҳ���ܷ����������á�
ͼ6c ʾAPC��Tϸ���ʵݿ�ԭ��ʱ����ϸ������������ϵ����m9-{, ��Ĵҽҩ
����Tϸ�����������ӻ��ƣ�����ΪTCR triggering(������TCR����)������һ�������ƣ����������ģʽ��һ�Ǿۼ�ģʽ(aggregation model)������TCRͬMHC/��ԭ�ĸ�������ʱ�����йط��ӵ������Ծۼ�(physical clustering)������������һ�ǹ��ͱ仯ģʽ(conformational change model)������TCRͬMHC/��ԭ�ĸ�������ʱ������ӽṹ�ĸı����������Davis����Ϊ[11]CD2����TCR����������һ���µĵ�3������ģʽ�����漰TCR�ľۼ����θı䣬ͼ6c����������һģʽ�ĺ��塣m9-{, ��Ĵҽҩ
���⣬CD45������Tϸ�����Ҳ��һ�������ã��������Ұ���������ø��(protein tyrosine phosphatase�� PTPase)������BCR�е����ã������Ѿ��ᵽ��ʵ��������ڴ����CD45-Tϸ��ϵͳ����TCR������ȫδ��ͬijЩ�ź�ͨ������ϵ(almost completely uncoupled)�������Ӷ�������ѧ���Ե��й��źţ������Ұ������ữ������������л������Ca[2+]ˮƽ������øC���IL-2���ڵȣ�����ijһCD45-Tϸ����TCR��Ӱ�������ֲ����������������ԡ�ԭ����CD45-/-С��Tϸ���ź�ͨ·ͬTCR�������ƵĽ�һ���о��Ľ��������CD45-/-Tϸ����TCRͬ�ź�ͨ·����ϵȱʧ(uncoupling)���в����Ե�(differentially)����һЩ�ź�ͨ·ͬTCR������������һЩ�ź�ͨ·ͬTCR�ֲ�����ȫ��������CD45������8��ͬ���칹��(isoforms)����ͬ��CD45ͬ���壬���в�ͬ������TCR�ź�ͨ·������������ȫ���ܵ��£����׳��ֵ�ì�ܣ������Ƕ�CD45����̬����ʶ������ѡ�m9-{, ��Ĵҽҩ
Frearson JA��������PTPaes��Tϸ����չ���������źŴ����е�����ʱ[12]����CD45�����൱��ϸ��������Tϸ���İ�����PTPaes����(cytosolic PTPase activities)����20��֮�࣬�е��Ѿ���¡����CD45����ĤPTPase���������ϱ�����CD45���õĵ�����P56lck��P59lynPTK(�����Ұ��ἤø)��������PTKs����Ϊ��ʹITAM���ữ�����ã�LTAM�Dz����� TCR����(invariant TCR polypeptide)��������У��������ữ����Ȼ�̼�TCR������ã�ITAM�����ữ��������RTK P56lck��P59lyn��ȥ���ữ��Frearson��ָ�����̼�TCR��һ��������P56lck��P59lyn��ȥ���ữ�����ܿ���������CD45�ı��������ά��ͬTCR������P59lyn��ͬCD4/CD8������P56lck�����ڸ�ˮƽ״̬��������ʹ����̼�ʱ�����źŵĴ��ݣ����ƺ�����TCR�̼���Ȼ��������������PTK��ȥ���ữ����CD45�ı���Ҳ��������ǵ�ȥ���ữ���ã��Ӷ��Tϸ�������߹��ܡ�Frearson����ͼ����ʽ����Tϸ����PTPase(Ĥ�ϣ����ڣ�����)�����Ϊͼ7�Ա�������ˡ�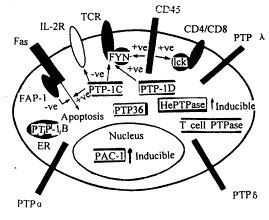
ͼ7 Tϸ����PTPase�ֲ�ʾ��*, ��Ĵҽҩ
ER��endoplasmic reticulum*, ��Ĵҽҩ
FAP-1��Fas-associated PTPase 1*, ��Ĵҽҩ
HePTPase��hematopoitic TPPase*, ��Ĵҽҩ
IL-2R��interleukin-2 receptor*, ��Ĵҽҩ
PTP-1C��proteintyrosine phosphotase 1C*, ��Ĵҽҩ
TCR��T cell receptor*, ��Ĵҽҩ
ͼ7ʾPTPø��Ĥ(CD45)��(PTP-1c PTP-1D��PTP38��PTP-1B��HePTPase��Tcell PTPase)��(PAC-1)�ķֲ���Ĥ�ϻ���PTP����PTP�ң��������˵ȣ���ͷʾ�йط��ӵĻ����ź���ϵ����FAP-1(Fas-associated PTPase 1)PTP-1C��PTP-1D�ȣ�PTPase����֮�࣬˵����Ե���������֮�ߣ�Ҳ����˵����ͬ��PTPase�����Ե������ڲ�ͬ�ĵ����ͼ7���ɿ�����Fap-1��Fas�Ļ������������ã���PTP-1C�����֮�дٽ����ã���Fas-1�Ĵٽ����ã�����ϸ��������ͬPTP-1C��TCR�ĸ���Ӧ��һ�µġ���PTP-1C�෴��PTP-1D������������TCR������ĤPTPase CD45��ȿ���������TCR-Fyn(P59)���ֿ���������TCR��Эͬ����CD4/CD8-Lck(P56)�������CD45����Tϸ���Ļ���������Ա����Dz���ȱ�ٵģ����ڱ���ǿ������CD45�����������Ժͺ�����PTPase���ӵ����ã��Ͳ��ٽ����ˡ�*, ��Ĵҽҩ
4 TCR��BCR���߹��ܱ���ķ�����̬�ԱȽ�*, ��Ĵҽҩ
TCR��BCR�ֱ���T��Bϸ���������߹��ܵ�������Ը�������ӣ�����ʶ��ԭ��ԭ�ĵ����á��Ƚ����ǵķ���ˮƽ����������������ߵ���̬�ԣ���Ȼ�������⣬Ҳ����˵�����߷�������ѧ�ѷ�չ�������߷�����̬ѧ�ij̶��ˡ�����������BCR�ṹ���ܵ���̬��ʱ��ָ����BCR������˫����ɼ���������ARH-1(����ԭʶ������)��ͼʾ�������ھ�ֹ̬�ͻ̬�ķ��ӹ��ͱ仯��CD45ʹIg����Fyn���ữ�Ŀ�����(ͼ2)����ͼʾ��BCR�ʱ���źŴ���������ϵ��CD22��CD19��CD21�ȷ���(ͼ3)����ʾ��BCR������̬�Եĸ����ԡ�������TCR�ṹ�빦�ܵ���̬��ʱ��ͼʾ��CD28��ϵ��PI3K(phosphoinositide 3 kinase)��ø�ź�ͬTCR�������ź�ͨ·֮���������ϵ(ͼ4)���ر�ͼʾ��Tϸ������IJ�ͬ������ӵķֲ���APCϸ����TCR�ʵݿ�ԭ��ʱ����ϸ������Ӵ������������������Ľ�Ϻͱ��������ϵ����������ǿ����CD2���ӵ���Ҫ����(ͼ6a��b��c)��
���⣬��BCR��п��������õķ���CD45��TCR�Ļ�о����������ɵ���Ҫ�ԣ�ͼ7�Դ˽�����ʾ��Ľ��͡�yx:, ��Ĵҽҩ
�Ƚ�TCR��BCR���ܱ������̬�ԣ��ص�����ָ����CDϵͳ������ӵ���ͬ��CD45��TCR��BCR�Ļ�ж������ã���ֻ��TCR�з������õ���CD4/CD8��CD2��CD28�ȣ�ֻ��BCR�з������õ���CD22��CD19��CD21�ȣ����д�����ʵ�����ϱ�������Щ���Ӳ���TCR��BCR��������Dz���ȱ�ٵġ��ɴ˿��Կ�����TCR��BCRʶ��ԭ�����ڷ���ˮƽ�ıȽϣ�ʵ�ʾ�������֮��ķ�����̬�ԵıȽϣ��������⡣yx:, ��Ĵҽҩ
��һ���ߣ��У�85�꣬��ѧ������yx:, ��Ĵҽҩ
5 ����yx:, ��Ĵҽҩ
��������ѧ��������ѧ�еĵ�λ����ȡ�ù㷺�Ĺ�ʶ��������̬ѧ��������ѧ�еĵ�λ����δ�ܵ��ձ��ע�⣬���ǽ����������IJ���������̬ѧ�Ĺ����Ժ�ѧ���ԣ��Ա�ʹ��һ�µ�ѧ�ƣ���ȡ��������ѧ�����ߵĹ�ʶ�������й����ۺ�ʵ���Ŀ��ع�������ϵ����������ѧ���Ͳ��ܲ����۷�����̬����ѧ�����⣬��������������TCR��BCR���ܱ�����µ����ϣ����Է�ӳ������̬����ѧ��������������ģ���ӭ���������ۡ�yx:, ��Ĵҽҩ
�ο�����yx:, ��Ĵҽҩ
1 �����.��������ѧ�ķ�չ.�����ѧ��̬��1964��10��7yx:, ��Ĵҽҩ
2 �������������.����ѧ�ľ����ԺͷǾ�����.����ѧ��־��1990��6��275yx:, ��Ĵҽҩ
3 �������������.������̬ѧ.������������.��̬ѧ.���������磬1988.46yx:, ��Ĵҽҩ
4 ������������أ�������.�����صķ�����̬ѧ.����ѧ��־��1993��9��71yx:, ��Ĵҽҩ
5 ��������������������أ���.���߷�����̬ѧ.����������̬ѧ���������������������������.�人��������ѧ���������磬1996yx:, ��Ĵҽҩ
6 Sonyang Z�� Shoelson SE��Chaudhuri M��et al. SH2 domains recognizeyx:, ��Ĵҽҩ
specific phosphopeptide sequenes. Cell��1993��72��767
7 Tseng J�� Lee YJ�� Eisfelder BJ��et al. The B cell antigen recptor complex�� mechanisms and implications of tyrosine kinases activation. Immunol Res��1994��13��299i!r8|\, http://www.100md.com
8 Pleiman CP��Clark MR�� Timson-Gauen LK��et al. Mapping of sites on the Src family protein tyrosine kinase P59 fyn and P56 lyn which interact with the effector molecules phospholipase C-gamma 2��microtubule-associated protein kinase��GTPase-activating protein�� and phosphatidylinositol 3-kinase. Mole Cell Biol��1993��13��5877i!r8|\, http://www.100md.com
9 Pezzatto A��Rabinovitch PS�� Dorkin B��et al.Role of the CD22 human B cell trigering by anti-immunoglobulin.J Immunnol��1988��140��1791i!r8|\, http://www.100md.com
10 Koretzky GA. The role of Grb2-associated proteins in T cell activation.i!r8|\, http://www.100md.com
Immunol Today��1997��18��401i!r8|\, http://www.100md.com
11 Davis SJ�� Vander Merwe PA. The structue and ligand interaction of CD2��implication for T cell function. Immunol Today��1996��17��177i!r8|\, http://www.100md.com
12 Frearson DA�� Alexander DR. Protein tyrosine phosphatases in T cell development�� apoptosis and signal. Immunol Today��1996��17��385i!r8|\, http://www.100md.com
(1997-12-04�ո�)(����� ��ѧ�� �� �� �� �� ������ ������)
��λ������ҽ�ƴ�ѧ����ѧ�о��� �人 430071w'(y/1, ��Ĵҽҩ
�ؼ��ʣ�w'(y/1, ��Ĵҽҩ
�� �� ѧ �� ־990301COMPARISON OF TCR-BCR ANTIGEN RECOGNITION MECHANISMw'(y/1, ��Ĵҽҩ
AT THE MOLECULAR ECOLOGICAL LEVELw'(y/1, ��Ĵҽҩ
��������ѧ��������ķ�չ�У������е��˱�����������ʵ���ˮƽ�ͳ���ˮƽ��������ı��ʣ�Ҳ�����������ڵĻ����������ṹ���ܣ�����ˮƽ�Թ⾵��ϸ��ѧ���漼��Ϊ���ڳ���ˮƽ�Ե羵�ͷ�������ѧ����Ϊ������ѧ��������ѧ�е�һ����Ҫ����һ��ѧ�Ƶķ�չ���������Ź���ĸı�Ͷ���������ʶ������ѧ�Բ����⡣�����Ƕ�����ѧ�ļ�ʶ�������ɼ�һ������ѧ����ϱ仯��չ���������ҹ���1964������˷�������ѧ�ĸ���[1]���ڵ�ʱ��˵�ǶԴ�ͳ����ѧ��һ�������Ըı䡣16���������������ѧ�����ԡ�����ѧ�ľ����ԺͷǾ����ԡ�Ϊ�⣬�Դ�ͳ������ѧ�������ߺͷ��������ߣ������������߸�����ᷨ�ϲ������������[2]���Ⱥ�õ���������һ������һ��Ȩ������ѧ��л���ĺ��������ȵ�֧�ֺ����͡��������ܵ�������̬ѧ��չ�������������ڹ����������������̬ѧ������[3]֮��������ȻҪ����̬ѧ�������뵽��������ѧ��������1993�������ˡ������صķ�����̬ѧ�������ķ���������ѧ��־������������ѧ��ժ(CA)ժ��[4]���ڿ����ǹ��������ġ�������̬ѧ����3ƪ�У���9��(16��24��)���������߷�����̬ѧ���й�����[5]������˵һ��ո�µ�����ѧ��֧ѧ��(������̬����ѧ)�Ѿ����������������Dz��϶����ķ�չ�����ʶ������̽�֡������Զ������û������кܶ��˽��TCR(Tϸ������)��BCR(Bϸ������)�Ĵ�ͳ������з�����̬ѧ�ķ����Ƚϣ����ѷ������߷�Ӧ�������غ�Эͬ����֮������ӹ�ϵ���ƺ����ģ�������ˣ����������������ӹ�ϵ����νģ���Խ������ۣ�����������ģ�������֮��w'(y/1, ��Ĵҽҩ
1 ʲô��T1Bϸ��ʶ����ԭ����������w'(y/1, ��Ĵҽҩ
T��Bϸ��ʶ��ԭ��������ӵ�Ȼ��TCR��BCR���������ڸ���ϸ���ı��档�����ǵ��������õ����֣������������������ͱ�����ӵ�Эͬ������CD����ϵͳ(Cluster desigation)�ȡ��������ֵĿ�ԭʶ�����������ܵ�MHC/��ԭ�ĸ�������Ϊ��������֮�⣬��Ҫ�ܵ�CD3��CD4��CD8�ȱ�����ӵ�Ӱ�죬CD3��TCR���ӵ���ɲ��֣�CD4��CD8�ֱ����Tϸ���Ĺ�������������ߵ�����(T����ϸ����Th)������ɱ�����ߵ�����(Tɱ��ϸ����Tk����ϸ����ϸ����TC)������һ����ΪTϸ����ϸ�����ߵ����壬������������ʶ��ԭ�ĵķ�����TCR���ⱻ��Ϊ����̼������ڷ��������߹���ʱ�����������CD4��CD8���ӵĴ̼�����ⱻ��ΪЭͬ�̼��������ǹ��ϵ�ϸ�����ߵķ��ӻ��ƣ�Ҳ��������ָ���������ӣ���ȻTCR������CD4��CD8�Ǵӡ��ӷ�����̬ѧ�ĸ�������������TCR��CD4��CD8��һ��(TCR-CD4��TCR-CD8)������Է��ӵĹ��ܿ���������Ϊ���ӷ����ǻ�Ϊ���������ģ�CD4��CD8����������Եı��֣�����������TCR�Ĵ��ڣ����������������Tϸ�����߷�Ӧ�е����þͲ����ܱ��ֳ�����TCRʶ�������Կ�ԭ�������ԣ�Ҳ����������CD4��CD8��Ϊ��������������Э��������TCR��������ϸ�����߹���Ҳ�����ܱ��ֳ�����������Dz���ð������������Է��ӵ���̬�Ե��¸���Ա���������̬ѧͬ��������ѧ��������֮�㣬�����ڷ�������ѧֻ��������Է��ӵ������ԣ�������̬ѧ��������Է��ӵ���̬�ԡ���̬�Ծ���ָ����ͬ������ͳһ���ں��ˮƽ������ĸ�����Ⱥ�壬�Ի����������������Ǿ��Եģ�����(ϸ��)ˮƽ�����ֹ�����ϸ���������书�ܻ��Ȼ���ض��Ļ����������ڳ���(����)ˮƽ�����ַ����������������Է��ӣ�Ҳ��Ȼ�����ض��ķ�����̬���������ǶԷ�����̬ѧ���������ᷨ���ɼ�Ϊ��������̬ѧ���о�������Է�����̬�ԵĿ�ѧ�����߷�����̬ѧ���о�������������Է�����̬�ԵĿ�ѧ����ϣ���������ۣ�����ӭ������
2 BCR�ṹ�빦�ܵ���̬��a&7n, ��Ĵҽҩ
Bϸ������Ȼ��ԭ��ʶ��ͷ�Ӧ�������ڱ������������ߵ�BCR��BCR�ܿ�ԭ�̼�������İ����ٲ��¼����ɵ���ϸ���Ļ�ͷֻ���ֱ������(anergy)����������ʼ����BCR�̼����Ե����Ұ�����øϵͳ���˼�ø������PLCr-1��2(��֬øCr-1��2)�����ữ�ͻ��ͬʱras����ͬShc��Grb2��mSOS����ɻ������(activation complex)���Ӷ�ʹRaf���ャĤ����˿/�հ�����ø��ٲ�������ϸ����ֳ�� ����Ӧ������߷�Ӧ��������Ҫ��a&7n, ��Ĵҽҩ
��ͳ�������������Һ���ߣ�ָ���ǿ�����ӣ����屾����λ��Bϸ��Ĥ�����������(Ig��immunoglobulin)���������Ե�һ���Ӵ��ڣ�ʵ����һ����˫����������ǵ���˫�壬��Ig����Ig�·ֱ���mb-1��B29������룬���߶��ǿ�Ĥ�ṹ������Ĥ��������Ĥ���Ͱ��������������ڵ�������(AA)����Ig��Ϊ61��Ig��Ϊ48AA��Ig����Ig�º��г�Ϊ��ԭʶ������(antigen recognition activation motif�� ARAM)��ƿ�ԭʶ��ͬ����-1(antigen recognition homology 1��ARH1)�������������ѷ����������ͼ1��ʾ��
a&7n, ��Ĵҽҩͼ1 Ig����Ig��AA���н�Ͽ�ԭ���ӱȽ�a&7n, ��Ĵҽҩ
����ʾAA�л���������ʾ������������Ҳ��֮�ı�a&7n, ��Ĵҽҩ
-��ʾ�����Բл�a&7n, ��Ĵҽҩ
ͼ1ʾBCR����������Ig����Ig�µ�AA���У�������ͬ��ӦЧӦ�������ϵ��������У�������������ARAM��ARH-1��Ig����Ig�¶�����ARH-1����ͬЧӦ�������ϵ������Ը��Բ�ͬ��Ig����DCSM���Ķ�ͬSrc���弤øLyn��Fyn��Blk��P38KDa��ϣ���Ig�º�QTAT���Ķ������������ֵ���40��42KDa��ͼ1����ʾ��ͨ��ת��������֤��Ig����DCSM��Ig�µ�QTAT�û�����Ig����ͬЧӦ���ӽ�������Ա�ΪIg���͵������ԣ���֮��Ȼ����Ig�����ԣ��б������[6]��YEGLN��YEDI�������ữʱ�������Ӷ�Fyn�Ľ����3���������߶������ữʱ������������20���������Ұ������ữ��ЭͬЧӦ(synergy effect)ָʾ��ЧӦ���ӽ�ϵĺ�����(co-operativity)��
Tseng��[7] �������ͷ���ģʽ�ر�������Ig�����ӹ��͵ľ�ֹ״̬��״̬����ʾ�ķ������������̬�Ա仯������ƣ�Tseng�������ģʽͼ��������Ϊͼ2�����Ա��ֳ�������̬�Եĺ��⣬��һģʽͼ��������Ϊͼ3������BCR������ʱ��Ϊ���ӵķ�����̬��������ϵ��
mq{&$, ��Ĵҽҩͼ2 BCR Ig���Ĺ��ͱ仯mq{&$, ��Ĵҽҩ
ͼ2ʾmIg BCR�İ���������˫��Ig����Ig�µĹ���ϵ����ʾ��ֹ̬����øFyn��N-�����ӽ��棬��ͬIg����DCSM��ϣ���SH2����ĩ��ø�������BCR�ܿ�ԭ�̼����Ig���������Ըı䣬��������CD45�������ã���Ϊ�Ұ�������ø(tyrosine phosphatase)���������������ã�һ�Ǵ�Fyn���۵������Ǵٽ�Fyn�ļ�ø���ԣ����ȶ���״̬��ͬʱʹIg����YEDI��YEGLNȥ���ữ�����⣬CD45ͨ��Ig��/Ig�����ữ��ȥ���ữ���̣����ܻ��дٽ��źŴ��ݵ����á�mq{&$, ��Ĵҽҩ
��һ���������Ѿ���ļ�ø��Fyn�ȼ���������BCR�������ϣ���Ϊͬ�����μ�ЧӦ����(secondary effectors)��ϵ����������磬Fyn��SH2�������Ig������SH3��������P13-k�����䰱�����ֽ����PLCr(phospholipase Cr)[8]������ͬIg������ϵ��FynͬP13-k��PLCr���ϵ���ø�������磬�������̬ϵ���临��Ҳ���ο�ͼ3��
mq{&$, ��Ĵҽҩͼ3 һ����BCR���帴����ģʽmq{&$, ��Ĵҽҩ
ͼ3ʾBCR�������Ӹ�������γɡ�ͬ�����ܷ�����ϵ��ЧӦ���ӣ�Ҳ��ͬ���������帴������ɰ�������(intracytoplasmic bridges)���źŴ��ݷ���Ӱ�졣����P13-k����ֻͬ����������Fyn��Lynͨ��SH3������ϵ����������SH2 ͬCD19���ӷ�����ϵ��ͼ3��ȷָ����BCR����������3���źŴ���;����������̼��������Ұ��ἤø�������Щ��ļ�ø������ʹIg����Ig�����ữ����ֱ�ӵĻ��ӵ�ʹCD19��CD22Ҳ���ữ������Щ���Ұ����Χ�����ǵ����ж�����ΪЧӦ������ʾ���ʹ������������Ĵμ��������źŴ��ݷ���(secondary and tertiary signal tranductors)�����á�����ʵ�������Bϸ������CD22�ı���Ͳ���ͨ��BCR�����ź�[9]����һ��Է�����̬����ѧ���ԣ��ر������壬��Ϊһ������ȱʧ(��һ������������ȱ��)�Ϳ�ʹ����Ӧ�еĹ��ܣ��������ܡ�
ͼ3�е�CD19��Ϊ�����ף�ͬCD21��ɸ����壬CD21���Dz�������2(CR2)����һ�����廹����TAPA���Ϸ��ӣ�CD19�ڽ���ΪSH2�ṩ������ϵ㣬����ͬ�������ӵ�������ϵ��h^9hcq, ��Ĵҽҩ
CD22Ϊ˫�壬��ΪCD22a��CD22b������������ͬԴ�����ԣ���3��ճ�Է��ӣ����走�ס���ϸ��ճ�Է��Ӻ�Ѫ��ճ�Է���(VCAM-1)��CD22a�鵼Bϸ��ճ���ڵ���ϸ���ͺ�ϸ����(��ϸ������)��CD22b����B-B��B-T֮����ճ�����á�h^9hcq, ��Ĵҽҩ
�����ǹ���BCR������̬�Եļ�Ҫ�����չ��δ�����ؽ����µĽ�չ����Ŀ�Դ���Ҳ��h^9hcq, ��Ĵҽҩ
3 TCR�ṹ�빦�ܵ���̬��h^9hcq, ��Ĵҽҩ
��ͳ��TCRʶ��ԭ�������Ի��ƣ�һֱ���DZȶ�BCR�ܵ��������ӵ����⣬���ﲻ������ϸ�����ۣ�ֻ�轫Koretzky[10]��Ward SG��[11]����Tϸ���������ʾ��ͼ���ۺ�Ϊ���ĵ�ͼ4���ٸ���˵�����ɣ������ǽ�ǿ��ijЩЭͬ���Ӷ�TCR���ܱ����е���Ҫ���壬����������BCR�Ľṹ����ʱ��Ҳ��ǿ������һ�㡣
h^9hcq, ��Ĵҽҩͼ4 TCR�Ľṹ�빦�ܼ�CD28������ʾ��h^9hcq, ��Ĵҽҩ
ERK��extracellular signal-regulated kinaseh^9hcq, ��Ĵҽҩ
FRAP��FK506-binding protein rapamycin associated proteinh^9hcq, ��Ĵҽҩ
JNK��c-jun N-terminal kinaseh^9hcq, ��Ĵҽҩ
PI3-kinase(PI3K)��phospoinositide 3-kinaseh^9hcq, ��Ĵҽҩ
PKC��protein kinase Ch^9hcq, ��Ĵҽҩ
PLC��phosphalipase Ch^9hcq, ��Ĵҽҩ
PtdIns(3��4��5)P3��phosphotidylinositol(3��4��5)-triphsphate
PTK��protein tryosine kinasdo, ��Ĵҽҩ
Ras��P21rasdo, ��Ĵҽҩ
SM��acidic sphingomeyelinasedo, ��Ĵҽҩ
TCR��T-cell receptordo, ��Ĵҽҩ
TOR��target of rapamycindo, ��Ĵҽҩ
TCRʵ���Ǧ�������ͬCD3 �ܦš��Ħż�˫�ɦ���ɵĸ����壬��δע�⸴���������ɵ���̬�����塣��ȻCD4��CD8����ΪTϸ����ͬ���ܱ��ֵı�Ƿ��ӣ��ܵ���һ���̶ȵ�ע�⣬Ҳ��δ�����ǿ��CD4��CD8��Tϸ���Ĺ��ܱ����ʵ���ž����Ե����ã�����������һ����Ϊ����CD4�����Tϸ���Dz�����ʹ������������õģ���CD�����Tϸ����Ҳ�Dz�����ʹ��ɱ�˰�ϸ�������õģ�����TCR�Ĺ��ܱ��֣�����ϵ��������CD8������CD28(ͼ4)��CD2�ȡ�TCR������Ҫ�źŴ���·���Ǧ�LC/Calcium P21ras�� PI3k�� P21ras��Ras����TCR�Ļ���CD28��P13k��ø���Ӷ�����CD28��TCR֮��ġ���̸��(cross talk)��P13k�����ܼ����ź����ε�Ras��Rac��Pkc��Bcl-XL�������P7056kҲ�����TCR���Ŀ��ͨ�����ӵ��ź�(���ǿɳ�֮Ϊ������̬����)�������ڻ�ʹٽ�IL-2�ı���Ӷ������ͼ�ǿTϸ���������ԡ�do, ��Ĵҽҩ
TCR�Ĺ��ܱ�����������ᵽ��CD28�⣬CD2�����ã�Ҳ���ܶ�֮����Ҫ��Ӱ�죬����Davis�ȶԴ������൱��ϸ�Ľ���[11]��CD2ʵ����һ������[����CD58��CD48��SLAM(T)��2B4(NK)��Ly-9(T2B)]����CEA(���߿�ԭ)����[ ����CD66c(NCA)��CD66b(CGMb)��CD66l(CEA)]ͬ��IgSF(�����׳�����)��ͼʾ����(ͼ5)���������ˡ�
do, ��Ĵҽҩͼ5 CD2��CEA��ΪIgSF�����������Աʾ��do, ��Ĵҽҩ
������Ĥ��
����glycosylphosphatidylinositol�� GPI��ʾí������l, ��Ĵҽҩ
TCRʶ��APC(antigen presentate cell)������ӳʵݵ�MHC-��ԭ��ʱ��CD2ͬAPC�ϵ�����CD58(LFA-3)�ܰ�ϸ�����ܷ���-3)��ϣ��м�ǿMHC���Ƶ�Tϸ��ʶ��ԭ�ĵ����á�Tϸ��ʶ��ԭ�Ĺؼ�������Tϸ����APC�ĽӴ�������֮���ж������͵ķ������ϣ���������ϸ���ϱ�����ӵ����Ͽ���Ϊ�ǻ�Ϊ����������Ľ�ϣ��м�ǿϸ����ϸ����ճ�����ã�ϸ��ճ�Է�����ϸ���������е���̬�Է���ֻ����ϸ�����߾����Ը����о������⣬�������߷�����̬ѧ�ľ������ݡ�Davis��������CD2������ʱ����ͼ����ʽ������ÿ�ַ��ӵ�ճ�����ã�����Ϊ����ͼ6a��b��c�����ʲο���
l, ��Ĵҽҩͼ6a ʾTϸ������ķ������Ƽ��������Сl, ��Ĵҽҩ
ITAM��immunoreceptor tyrosine based activation motifl, ��Ĵҽҩ
GPI����ͼ5l, ��Ĵҽҩ
����Ҫָ�����ǣ�TCRCD3�����庬�����������Ұ�������(ITAM)���ڽ�CD2���ӣ�CD2��TCR��������ݽᾧѧ�͵羵�����о�������ʾ����CD45��CD43ҪС6��8���������ǵı����ܶ��ֱ�TCR��CD2��2��3��(ͼ6a��b)
l, ��Ĵҽҩͼ6b ʾ��ֹ��Tϸ���ı�����ӣ��ص�������ʾCD2��CD45��CD4��TCRCD3�ȵķֲ���ϵl, ��Ĵҽҩ
CD2ͬABC��������ʱ���ɵ���T��APC��ϸ��Ĥ���ܽӴ�������CD45�ķ��ӽϴ����Ե�TCRͬMHC/��ԭ�ĺ�CD2ͬ������ʱ�����뽫CD45�������Ա��ſ�������������ϸ������Ӵ�����ͼ6c���Կ���CD2/����ͬTCR/MHC��ԭ�ĸ�������ɸ����ӵĸ�����ϵͳ���縴�����������������������CD4���Ӳ�ͬLck��ø����ϵ��ֻ�и�������TCR��ʶ��ԭ�ģ��Ӷ�ʹLck����ITAM���ữ���ã�����һ�����ZAP-70���������ữ����źţ�����TCR���������Եijɹ��������Ӧ�ñ�����IL-2�Ļ�����IL-2�����ϡ������������CD4ȱʧ��MHCδ�п�ԭ�Ľ�ϣ���Ȼ���ᷢ���������ã������������CD4�Ϳ�ԭ�Ĵ��ڣ�����CD2/����IJ��룬Ҳ���ܷ����������á�
ͼ6c ʾAPC��Tϸ���ʵݿ�ԭ��ʱ����ϸ������������ϵ����m9-{, ��Ĵҽҩ
����Tϸ�����������ӻ��ƣ�����ΪTCR triggering(������TCR����)������һ�������ƣ����������ģʽ��һ�Ǿۼ�ģʽ(aggregation model)������TCRͬMHC/��ԭ�ĸ�������ʱ�����йط��ӵ������Ծۼ�(physical clustering)������������һ�ǹ��ͱ仯ģʽ(conformational change model)������TCRͬMHC/��ԭ�ĸ�������ʱ������ӽṹ�ĸı����������Davis����Ϊ[11]CD2����TCR����������һ���µĵ�3������ģʽ�����漰TCR�ľۼ����θı䣬ͼ6c����������һģʽ�ĺ��塣m9-{, ��Ĵҽҩ
���⣬CD45������Tϸ�����Ҳ��һ�������ã��������Ұ���������ø��(protein tyrosine phosphatase�� PTPase)������BCR�е����ã������Ѿ��ᵽ��ʵ��������ڴ����CD45-Tϸ��ϵͳ����TCR������ȫδ��ͬijЩ�ź�ͨ������ϵ(almost completely uncoupled)�������Ӷ�������ѧ���Ե��й��źţ������Ұ������ữ������������л������Ca[2+]ˮƽ������øC���IL-2���ڵȣ�����ijһCD45-Tϸ����TCR��Ӱ�������ֲ����������������ԡ�ԭ����CD45-/-С��Tϸ���ź�ͨ·ͬTCR�������ƵĽ�һ���о��Ľ��������CD45-/-Tϸ����TCRͬ�ź�ͨ·����ϵȱʧ(uncoupling)���в����Ե�(differentially)����һЩ�ź�ͨ·ͬTCR������������һЩ�ź�ͨ·ͬTCR�ֲ�����ȫ��������CD45������8��ͬ���칹��(isoforms)����ͬ��CD45ͬ���壬���в�ͬ������TCR�ź�ͨ·������������ȫ���ܵ��£����׳��ֵ�ì�ܣ������Ƕ�CD45����̬����ʶ������ѡ�m9-{, ��Ĵҽҩ
Frearson JA��������PTPaes��Tϸ����չ���������źŴ����е�����ʱ[12]����CD45�����൱��ϸ��������Tϸ���İ�����PTPaes����(cytosolic PTPase activities)����20��֮�࣬�е��Ѿ���¡����CD45����ĤPTPase���������ϱ�����CD45���õĵ�����P56lck��P59lynPTK(�����Ұ��ἤø)��������PTKs����Ϊ��ʹITAM���ữ�����ã�LTAM�Dz����� TCR����(invariant TCR polypeptide)��������У��������ữ����Ȼ�̼�TCR������ã�ITAM�����ữ��������RTK P56lck��P59lyn��ȥ���ữ��Frearson��ָ�����̼�TCR��һ��������P56lck��P59lyn��ȥ���ữ�����ܿ���������CD45�ı��������ά��ͬTCR������P59lyn��ͬCD4/CD8������P56lck�����ڸ�ˮƽ״̬��������ʹ����̼�ʱ�����źŵĴ��ݣ����ƺ�����TCR�̼���Ȼ��������������PTK��ȥ���ữ����CD45�ı���Ҳ��������ǵ�ȥ���ữ���ã��Ӷ��Tϸ�������߹��ܡ�Frearson����ͼ����ʽ����Tϸ����PTPase(Ĥ�ϣ����ڣ�����)�����Ϊͼ7�Ա�������ˡ�
ͼ7 Tϸ����PTPase�ֲ�ʾ��*, ��Ĵҽҩ
ER��endoplasmic reticulum*, ��Ĵҽҩ
FAP-1��Fas-associated PTPase 1*, ��Ĵҽҩ
HePTPase��hematopoitic TPPase*, ��Ĵҽҩ
IL-2R��interleukin-2 receptor*, ��Ĵҽҩ
PTP-1C��proteintyrosine phosphotase 1C*, ��Ĵҽҩ
TCR��T cell receptor*, ��Ĵҽҩ
ͼ7ʾPTPø��Ĥ(CD45)��(PTP-1c PTP-1D��PTP38��PTP-1B��HePTPase��Tcell PTPase)��(PAC-1)�ķֲ���Ĥ�ϻ���PTP����PTP�ң��������˵ȣ���ͷʾ�йط��ӵĻ����ź���ϵ����FAP-1(Fas-associated PTPase 1)PTP-1C��PTP-1D�ȣ�PTPase����֮�࣬˵����Ե���������֮�ߣ�Ҳ����˵����ͬ��PTPase�����Ե������ڲ�ͬ�ĵ����ͼ7���ɿ�����Fap-1��Fas�Ļ������������ã���PTP-1C�����֮�дٽ����ã���Fas-1�Ĵٽ����ã�����ϸ��������ͬPTP-1C��TCR�ĸ���Ӧ��һ�µġ���PTP-1C�෴��PTP-1D������������TCR������ĤPTPase CD45��ȿ���������TCR-Fyn(P59)���ֿ���������TCR��Эͬ����CD4/CD8-Lck(P56)�������CD45����Tϸ���Ļ���������Ա����Dz���ȱ�ٵģ����ڱ���ǿ������CD45�����������Ժͺ�����PTPase���ӵ����ã��Ͳ��ٽ����ˡ�*, ��Ĵҽҩ
4 TCR��BCR���߹��ܱ���ķ�����̬�ԱȽ�*, ��Ĵҽҩ
TCR��BCR�ֱ���T��Bϸ���������߹��ܵ�������Ը�������ӣ�����ʶ��ԭ��ԭ�ĵ����á��Ƚ����ǵķ���ˮƽ����������������ߵ���̬�ԣ���Ȼ�������⣬Ҳ����˵�����߷�������ѧ�ѷ�չ�������߷�����̬ѧ�ij̶��ˡ�����������BCR�ṹ���ܵ���̬��ʱ��ָ����BCR������˫����ɼ���������ARH-1(����ԭʶ������)��ͼʾ�������ھ�ֹ̬�ͻ̬�ķ��ӹ��ͱ仯��CD45ʹIg����Fyn���ữ�Ŀ�����(ͼ2)����ͼʾ��BCR�ʱ���źŴ���������ϵ��CD22��CD19��CD21�ȷ���(ͼ3)����ʾ��BCR������̬�Եĸ����ԡ�������TCR�ṹ�빦�ܵ���̬��ʱ��ͼʾ��CD28��ϵ��PI3K(phosphoinositide 3 kinase)��ø�ź�ͬTCR�������ź�ͨ·֮���������ϵ(ͼ4)���ر�ͼʾ��Tϸ������IJ�ͬ������ӵķֲ���APCϸ����TCR�ʵݿ�ԭ��ʱ����ϸ������Ӵ������������������Ľ�Ϻͱ��������ϵ����������ǿ����CD2���ӵ���Ҫ����(ͼ6a��b��c)��
���⣬��BCR��п��������õķ���CD45��TCR�Ļ�о����������ɵ���Ҫ�ԣ�ͼ7�Դ˽�����ʾ��Ľ��͡�yx:, ��Ĵҽҩ
�Ƚ�TCR��BCR���ܱ������̬�ԣ��ص�����ָ����CDϵͳ������ӵ���ͬ��CD45��TCR��BCR�Ļ�ж������ã���ֻ��TCR�з������õ���CD4/CD8��CD2��CD28�ȣ�ֻ��BCR�з������õ���CD22��CD19��CD21�ȣ����д�����ʵ�����ϱ�������Щ���Ӳ���TCR��BCR��������Dz���ȱ�ٵġ��ɴ˿��Կ�����TCR��BCRʶ��ԭ�����ڷ���ˮƽ�ıȽϣ�ʵ�ʾ�������֮��ķ�����̬�ԵıȽϣ��������⡣yx:, ��Ĵҽҩ
��һ���ߣ��У�85�꣬��ѧ������yx:, ��Ĵҽҩ
5 ����yx:, ��Ĵҽҩ
��������ѧ��������ѧ�еĵ�λ����ȡ�ù㷺�Ĺ�ʶ��������̬ѧ��������ѧ�еĵ�λ����δ�ܵ��ձ��ע�⣬���ǽ����������IJ���������̬ѧ�Ĺ����Ժ�ѧ���ԣ��Ա�ʹ��һ�µ�ѧ�ƣ���ȡ��������ѧ�����ߵĹ�ʶ�������й����ۺ�ʵ���Ŀ��ع�������ϵ����������ѧ���Ͳ��ܲ����۷�����̬����ѧ�����⣬��������������TCR��BCR���ܱ�����µ����ϣ����Է�ӳ������̬����ѧ��������������ģ���ӭ���������ۡ�yx:, ��Ĵҽҩ
�ο�����yx:, ��Ĵҽҩ
1 �����.��������ѧ�ķ�չ.�����ѧ��̬��1964��10��7yx:, ��Ĵҽҩ
2 �������������.����ѧ�ľ����ԺͷǾ�����.����ѧ��־��1990��6��275yx:, ��Ĵҽҩ
3 �������������.������̬ѧ.������������.��̬ѧ.���������磬1988.46yx:, ��Ĵҽҩ
4 ������������أ�������.�����صķ�����̬ѧ.����ѧ��־��1993��9��71yx:, ��Ĵҽҩ
5 ��������������������أ���.���߷�����̬ѧ.����������̬ѧ���������������������������.�人��������ѧ���������磬1996yx:, ��Ĵҽҩ
6 Sonyang Z�� Shoelson SE��Chaudhuri M��et al. SH2 domains recognizeyx:, ��Ĵҽҩ
specific phosphopeptide sequenes. Cell��1993��72��767
7 Tseng J�� Lee YJ�� Eisfelder BJ��et al. The B cell antigen recptor complex�� mechanisms and implications of tyrosine kinases activation. Immunol Res��1994��13��299i!r8|\, http://www.100md.com
8 Pleiman CP��Clark MR�� Timson-Gauen LK��et al. Mapping of sites on the Src family protein tyrosine kinase P59 fyn and P56 lyn which interact with the effector molecules phospholipase C-gamma 2��microtubule-associated protein kinase��GTPase-activating protein�� and phosphatidylinositol 3-kinase. Mole Cell Biol��1993��13��5877i!r8|\, http://www.100md.com
9 Pezzatto A��Rabinovitch PS�� Dorkin B��et al.Role of the CD22 human B cell trigering by anti-immunoglobulin.J Immunnol��1988��140��1791i!r8|\, http://www.100md.com
10 Koretzky GA. The role of Grb2-associated proteins in T cell activation.i!r8|\, http://www.100md.com
Immunol Today��1997��18��401i!r8|\, http://www.100md.com
11 Davis SJ�� Vander Merwe PA. The structue and ligand interaction of CD2��implication for T cell function. Immunol Today��1996��17��177i!r8|\, http://www.100md.com
12 Frearson DA�� Alexander DR. Protein tyrosine phosphatases in T cell development�� apoptosis and signal. Immunol Today��1996��17��385i!r8|\, http://www.100md.com
(1997-12-04�ո�)(����� ��ѧ�� �� �� �� �� ������ ������)