EB病毒LMP1在鼻咽癌细胞中调控核转录因子κB活性研究
作者:廖伟 唐敏 邓锡云 曹亚
单位:湖南医科大学肿瘤研究所,湖南 长沙 410078
关键词:潜伏膜蛋白1(LMP1);核转录因子κB;鼻咽癌
病毒学报000302 摘要:为了探讨EB病毒潜伏膜蛋白1(LMP1)的致瘤机制,对鼻咽癌细胞中LMP1通过核转录因子κB(NFκB)介导的信号传导途径在鼻咽癌变中的意义进行了研究。利用LMP1受四环素衍生物强力霉素诱导表达的鼻咽癌细胞Tet-on-LMP1 HNE2,通过NFκB报道基因分析法、凝胶迁移率分析(EMSA)及细胞集落形成率等方法,结合硫代磷酸反义寡核苷酸阻断技术,证实LMP1增强鼻咽癌细胞NFκB的DNA结合活性和反式激活活性,这种增强的活性可被LMP1及NFκB p65硫代磷酸反义寡核苷酸阻断,而NFκB p50仅阻断DNA结合活性,对反式激活活性无影响。同时,LMP1及NFκB p65硫代磷酸反义寡核苷酸可部分逆转鼻咽癌细胞的恶性表型,而反义p50这种效应不显著。这些结果表明,LMP1在鼻咽癌中通过NFκB信号传导途径可能参与了鼻咽癌变,NFκB p65可能是主要的效应者。
, 百拇医药
中图分类号:R373 文献标识码:A
文章编号:1000-8721(2000)03-0198-05
The Epstein-Barr Virus Encoded Latent Membrane Protein 1 Promotes Nuclear Factor-kappa B Activity in a Nasopharyngeal Carcinoma Cell Line
LIAO Wei,TANG Min,DENG Xi-yun,CAO Ya
(Cancer Research Institute, Hunan Medical University, Changsha, Hunan 410078, China)
Abstract: As an oncogenic protein, the Epstein-Barr virus encoded latent membrane protein 1(LMP1)may play a key role in nasopharyngeal carcinogenesis through its signal transduction pathway, especially NF-κB pathway. In order to attest this hypothesis, a dual-stable LMP1 integrated nasopharyngeal carcinoma cell line with Tet-on regulating system designated Tet-on-LMP1 HNE2, in which expression of LMP1 could be dose-dependent regulated by doxycycline, was used. It was found that NFκB activity was 3-10 folds high at LMP1 induced statement by reporter gene analysis, and this result was further confirmed by electrophoresis mobility shift assay which showed NFκB accumulated in nuclear when LMP1 expression was induced. When Tet-on-LMP1 HNE2 cells were co-incubated with phosphorothioate oligonucleotides (PS-ODNs) to antisense LMP1, NFκB p65 and NFκB p50 at various times, NFκB activity curves showed that antisense LMP1 blocked NFκB activity at earlier time than antisense p65 did, and p50 showed no block effect. What's more, PS-ODNs to antisense LMP1 and NFκB p65, but not p50, partially reversed malignant phenotype of Tet-on-LMP1 HNE2 boosted by LMP1 expression. All of these deduced that LMP1 could participate in the nasopharyngeal carcinogenesis by improving NFκB activity in NPC, and NFκB, of which p65 in particular, has the potency used as a molecular target to NPC gene therapy.
, 百拇医药
Key words: Epstein-Barr virus latent membrane protein 1; nuclear factor-kappa B(NFκB); nasopharyngeal carcinoma
核转录因子κB(NFκB)是近年来研究的热点,因与免疫球蛋白kappa轻链基因(Igκ基因)的3′末端增强子结合而得名[1]。NFκB属于一个高度保守的反式激活因子家族,由多个成员组成,包括Dif、Dorsal、Relish、v-rel、c-rel、p50(NF-κB1)、p65(RelA)、RelB和p52。家族中每个成员都包含N末端由300个氨基酸组成的保守序列,称rel同源区域(rel homology domain,RHD)。该区域负责DNA-binding、二聚体聚合及同IκB家族结合等功能,其中Dif、Dorsal、Relish、v-rel、c-rel、p65(RelA)和RelB C末端具有与反式激活相关的部位(transactivation site),p52与p50则没有。NFκB家族成员之间可形成同源二聚体如p50∶p50,或异源二聚体如p50∶p65等。在大多数细胞类型中,NFκB表现为p50∶p65二聚体形式,其他类型的二聚体同时也有低水平的表达[2]。
, 百拇医药
位于胞浆内的NFκB一般与NFκB活性抑制因子IκBα结合,其Rel同源区域被遮蔽,从而不能进入核内发挥反式激活活性。在外来刺激因素,如UV紫外线、氧自由基、病毒和细菌内毒素等的作用下,IκBα被磷酸化并降解,NFκB与IκBα解离,由胞浆转位入细胞核,启动具有κB结合位点的基因转录表达。作为重要的转录调控因子,NFκB调节很多的基因表达,这些基因与细胞增殖、分化、免疫反应、凋亡、转化有着密切关系[3]。
EB病毒是已被认为与鼻咽癌发病密切相关的因素,其编码的潜伏膜蛋白LMP1由于具有潜在的致瘤作用,而成为近年来EB病毒致瘤机制研究的热点。研究发现,LMP1可能类似于肿瘤坏死因子家族成员CD40,在细胞增殖及凋亡的信号传导途径中具有重要的作用。在LMP1所介导的细胞调控网络系统中,NFκB处于信号传导枢纽。近年来研究发现,LMP1蛋白至少可以通过两条途径磷酸化IκBα并活化NFκB:一条为肿瘤坏死因子相关蛋白(tumor necrosis factor receptor associated factors,TRAFs)途径[4];另一条为肿瘤坏死因子受体死亡结构域(tumor necrosis factor receptor associated death domain protein,TRADD)途径[5]。其中TRAFs与CTAR1结合,激活大约25%的NFκB活性;TRADD与CTAR2结合,75%的NFκB的活化与此相关。当LMP1的6个跨膜结构域聚合后,TRAFs和TRADD与LMP1上各自的结构域结合,激活Iκ激酶,Iκ激酶活化后,促使IκBα磷酸化,活化NFκB。
, 百拇医药
作为与鼻咽癌发病密切相关的EB病毒瘤基因,LMP1能通过磷酸化IκBα而活化NFκB,从而提示我们探讨在鼻咽癌中LMP1能否调节NFκB的活性及LMP1是否通过NFκB信号传导途径参与鼻咽癌变的过程等问题,对于阐述EB病毒致鼻咽癌的可能机理并提出鼻咽癌“靶分子”治疗新策略具有较为重要的意义。
材料与方法
1 细胞系 鼻咽癌细胞系HNE2是湖南医科大学肿瘤研究所建立的EB病毒阴性的低分化鳞癌细胞株[6]。PCR进一步证实其中LMP1为阴性。经两轮稳定的转染,将四环素基因调控系统导入其中,构建LMP1受四环素衍生物强力霉素诱导表达的鼻咽癌细胞系pTet-on-LMP1 HNE2[7]。细胞培养液为含15%小牛血清的RPMI-1640。37℃ 5%CO2维持200mg/L G418和50mg/L潮霉素以筛选药物浓度。
2 硫代磷酸化寡脱氧核苷酸有义和反义NFκB p65及p50的设计 硫代磷酸化修饰的寡脱氧核苷酸是进行反义核酸干预的有效方法。根据文献[8],设计针对p65及p50的3′末端硫代磷酸化修饰的有义和反义寡脱氧核苷酸,NFκB p65有义寡脱氧核苷酸:5′-CGGCCATGGACGAACTGTTC-3′,反义寡脱氧核苷酸:5′-GAACAGTTCGTCCACATGGCCCA-CTTC-3';NFκBp50有义寡脱氧核苷酸;5'-ATCTTCACCATGGCAGCAGA-3',反义寡脱氧核苷酸:5'-ATCGTCTGCCATGGTGAAGAT-3'。以上寡脱氧核苷酸由美国国立癌症研究院DNA重组实验室合成,使用浓度为20mg/L浓度。
, 百拇医药
3 NFκB报道基因分析 用NFκB报道基因质粒pNFκB-luc的荧光素酶表达分析NFκB的活性。pNFκB-luc包含螟蛾科荧光素酶基因,它的表达受CMV启动子调控,在CMV启动子中插入了来源于HIV LTR的κB元件。用LipofecTAMINE的方法将pNFκB-luc一过性转染入细胞,转染12~48h后,按荧光素酶分析系统的说明,加入100μl 1×裂解缓冲液,室温下裂解15min,用细胞刮子将细胞刮下,吸至离心管内,12?000r/min离心,取20μl上清液,加入100μl反应底物(Luciferaly-CoA),迅速推入液闪仪(BECKMAN LS5000 TA)单光子档,检测cpm值。间隔10s,每一个样本读两个数值,取均值。
4 凝胶迁移率分析(electrophoresis mobility shift assay,EMSA) 细胞核蛋白的提取参照Ichikawa[9]等的方法进行,收集细胞,冷PBS洗1次,约107个细胞用1ml SMTD-PMSF(0.25mol/L蔗糖,1mmol/L MgCl2,20mmol/L Tris-HCl pH7.85,1mmol/L DTT,0.1mmol/LPMSF)重悬细胞,玻璃匀浆器匀浆4次,加入Triton X-100至终浓度1%,再匀浆2次,离心(1?000g×10min),弃上清,1ml含0.5%Triton X-100的SMTD-PMSF重悬沉淀,玻璃匀浆器再匀浆2次,离心,沉淀再用1ml含1mmol/L CaCl2的SMTD-PMSF洗1次,离心,用与细胞核等体积的KMTD(0.3%KCl,1mmol/L,MgCl2,10mmol/L Tris-HCl pH8.0,1mmol/L DTT)室温温育1h,每隔15min剧烈振动1次,15?000g×15min离心,保留上清核提取物。蛋白浓度用BCA检测试剂(Pierce Chemical Co,Rockford,IL)测定。EMSA分析按照Promega公司Gel shift assay kit操作手册的说明进行。首先用γ-32P-ATP在T4多核苷酸激酶的作用下,连接至NFκB寡核苷酸,37℃ 20min EDTA终止反应,加入89μl TE缓冲液,用G-25柱层析方法纯化探针。约5μg核蛋白与Kit中的结合缓冲液(binding buffer)作用10min,同时以Kit中提供的HeLa细胞核蛋白10μg作为阳性对照;加入1μl纯化后的32P标记NFκB consensus oligo,室温放置约3h,加入1μl加样缓冲液,于4%聚丙烯酰胺凝胶(acrylamide∶bis为39∶1)进行电泳分离。待加样缓冲液中的染料溴酚蓝距电泳槽底部1/4距离时,终止电泳。用带有增感屏的暗闸进行X光片显影、曝光。
, 百拇医药
5 细胞集落形成分析 配制0.5%的琼脂,以该琼脂为底层,将dox(+)、dox(-)的Tet-on-LMP1 HNE2、以20μg/ml正义或反义LMP1及p65、p50处理24h后的Tet-on-LMP1 HNE2 2×104个细胞加入培养基,再用此培养基稀释0.5%的琼脂至0.33%,作为上层琼脂。每组种3个平皿,置5%CO2 37℃培育10天,显微镜下用随机取样法观察每一平皿5个1cm2内克隆数目(8个细胞以上为一集落)及大小。
结 果
1 LMP1促进NFκB的反式激活活性将Tet-on-LMP1 HNE2细胞在dox(-)、dox(+)及分别加入硫代磷酸化修饰的反义或正义NFκB p65的状态下培养36h,用NFκB报道质粒检测NFκB的反式激活活性。由图1可看到,在dox(+)状态或dox(+)状态加上正义NFκB p65,NFκB具有较高的反式激活活性;dox(-)或dox(+)加上反义NFκB p65,则NFκB反式激活活性下降;作为对照,未转染Tet-on调控系统的HNE2即使在dox(+)状态,NFκB反式激活活性也较低。这说明LMP1可促进NFκB的反式激活活性,这种作用可被反义NFκB p65所阻断。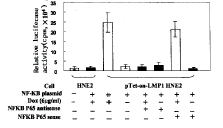
, 百拇医药
图1 LMP1增强NFκB的反式激活活性
Figure 1 LMP1 increased transactivational activity of NFκB
Tet-on-LMP1 HNE2 cell with or without dox, or coincubated with sense and antisense NFκB p65 for 36h was analyzed by means of NFκB reporter gene to detect NFκB transactivational activity. This activity was higher in cells with dox(+), in which expression of LMP1 was induced, and could be blocked by antisense NFκB p65.
, 百拇医药 2 LMP1对NFκB活性调节的时间效应
在证实LMP1促进NFκB反式激活活性的基础上,为了进一步阐明LMP1对NFκB反式激活活性调节的时间效应,并观察与p65和p50对NFκB反式激活活性的影响的差别,以PS-ODNs反义LMP1,p65和p50 20μg/ml与诱导和非诱导状态的Tet-on-LMP1 HNE2细胞共温育,并检测温育后12h、24h、36h和48hNFκB活性,可以看到,反义LMP1导入后,在所有的时间点NFκB活性都较低(图2A);反义p65 PS-ODNs则在导入后36h才产生抑制效应,至48h活性有所回升(图2B)。而反义p50 PS-ODNs在48h内对NFκB的活性几乎都不产生影响(图2C)。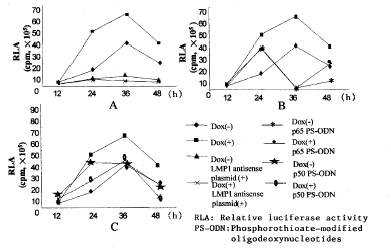
图2 LMP1对NFκB反式激活活性调节的时间效应
Figure 2 Time-course of LMP1 mediated NFκB transactivational activity
, 百拇医药
(A)Antisense LMP1 blocked the NFκB activity. 2μg/ml LMP1 antisense plasmid was transiently transfected into pTet-on-LMP1 HNE2 cell line with or without 2μg/ml dox. Markedly blocked effect was shown at least 12h after pNFκB-luc transfection. (B)Antisense P65 oligonucleotide blocked the NFκB activity. 20μg/ml P65 antisense oligonucleotide was added to the medium of pTet-on-LMP1 cells with or without 2μg/ml dox at indicated time. The most distinct blocked effect was shown at 36h after pNFκB-luc transfection. However, p50 antisense oligonucleotide didn't block the NFκB transactivational activity(C).
, http://www.100md.com
3 LMP1、NFκB p65和NFκB p50调节NFκB的DNA结合活性
LMP1、NFκB p65可以调控NFκB的反式激活活性,而NFκB p50则无这种作用。而这3个因子对NFκB的DNA结合活性是否有影响,利用凝胶移动迁移率(EMSA)进行了进一步探讨。EMSA的结果显示,HeLa阳性对照呈现明显的NFκB两条信号,而对于Tet-on-LMP1 HNE2细胞,在dox(+)状态下,NFκB的DNA结合活性增强,加入硫代磷酸化修饰的反义(antisense)LMP1,p65和p50共温育36h后,这种结合活性显著下降(图3)。
4 LMP1和NFκB p65和NFκB p50对鼻咽癌恶性表型的影响
倒置显微镜下观察dox(+)、dox(-)的Tet-on-LMP1 HNE2,dox(+)加上20μg/ml正义或反义LMP1和dox(+)加p65、p50处理36h后的Tet-on-LMP1 HNE2的细胞集落形成情况,可以看到,在dox(+)状态下细胞集落最为密集,且集落体积较大;在dox(-)、反义LMP1及p65处理后,则细胞集落明显稀疏,体积较小,而反义p50则作用相对不明显(图4)。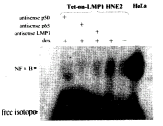
, 百拇医药
图3 LMP1对NFκB结合活性的调节
Figure 3 NFκB binding activity regulated by LMP1
NFκB binding activity blocked by antisense LMP1, antisense p65 and antisense p50. NFκB binding activity in Tet-on-LMP1 HNE2 cell line following dox, antisense LMP1, antisense p65 and antisense p50 treatment was analyzed by means of EMSA. Tet-on-LMP1 HNE2 cell line was plated in ten 100-mm cell culture dishes(1×106 cells/dish)in 1640 with 10% fetal bovine serum(FBS)and cultured for 4h. The cell medium was replaced by 1640 with 10% FBS and with dox, antisense LMP1, antisense p65 and antisense p50 and cultured for 36h. 10μg of nuclear protein purified from cells were incubated with 3×104 cpm 32P-labeled consensus NFκB oligonucleotide and separated by 5% acrylamide gel electrophoresis as described in “Materials and Methods”.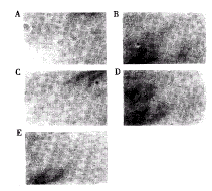
, 百拇医药
图4 细胞集落形成检测LMP1、p65和p50对Tet-on-LMP1 HNE2鼻咽癌细胞系恶性表型的影响
Figure 4 Colony forming of Tet-on-LMP1 HNE2 cell line treated as conditions indicated
A. dox(+); B. dox(-); C. dox(+)with PS-ODN LMP1 treated; D. dox(+)with p65 PS-ODN LMP1 treated; E. dox(+)with PS-ODN p50 treated. Experimental procedure was described in detail in "Materials and Methods".
讨 论
LMP1的致瘤机制近年来一直是一个研究的热点。Kieff[10]于1996年提出,LMP1可能类似于肿瘤坏死因子受体家族成员CD40,通过参与细胞内信号传导网络,而参与细胞增殖或凋亡的调控。目前已知LMP1至少与3条信号传导通路有关:①通过TRAFs/TRADD活化NFκB;②通过SEK-JNK1及Ras-Rafl-MER-Erk1/2活化AP-1;③通过JAK-STAT途径调节相关基因的表达。其中LMP1通过NFκB调控其靶基因的表达尤为重要。
, 百拇医药
NFκB的活化可能与许多肿瘤的发生有关。将v-Rel置于T细胞特异性启动子的控制下产生的转基因小鼠会发生T细胞淋巴瘤。此外,NFκB可分别与HTLV-1逆反式激活病毒和HBV病毒中的Tax及x基因产物结合,在它们的致瘤中产生作用。在人T细胞和B细胞肿瘤中大约有1%~10%的Rel或IκBα家族基因发生改变。人乳腺癌和多发性骨髓瘤中均可见NFκB的点突变活化[3]。
本工作利用一株LMP1受四环素衍生物强力霉素诱导表达的鼻咽癌细胞株Tet-on-LMP1 HNE2,探讨了LMP1在鼻咽癌细胞中调控NFκB的活性,并通过这种活性的调控,对鼻咽癌细胞恶性表型产生的影响。
NFκB的活性包括DNA结合活性和反式激活活性两种。用NFκB报道基因分析法可检测其反式激活活性,而凝胶迁移率分析显示的为其DNA结合活性。研究显示,尽管LMP1、p65和p50的反义核苷酸都降低NFκB的DNA结合活性,但三者对其反式激活活性的影响是不同的。LMP1可促进NFκB的反式激活活性,这种作用被反义NFκB p65所阻断;反义LMP1导入后,NFκB的活性在所有的时间点都较低;反义p65 PS-ODNs则在导入后36h才产生抑制效应,而48h活性有所回升。而反义p50 PS-ODNs在48h内对NFκB的反式激活活性几乎都不产生影响。这说明LMP1通过磷酸化IκBα,对已存在于胞浆内的NFκB的活化较早期发生,而反义p65对NFκB活性的影响,可能是先通过抑制p65 RNA合成,再降低p65蛋白的翻译,最终影响NFκB的活性,反义p50的干预对NFκB活性不产生作用,进一步提示p50对NFκB的反式激活活性无影响。
, http://www.100md.com
细胞集落形成实验证实LMP1可促进细胞集落形成率,而反义LMP1和反义p65可部分阻断这种恶性表型,反义p50则作用不显著。
由此我们推断,EB病毒LMP1通过活化NFκB,继而调节相关基因的表达,在鼻咽癌的癌变过程中可能发挥了重要作用。其中,反义p65寡核苷酸对NFκB反式激活活性的阻断及对细胞集落形成的影响,提示应进一步探讨鼻咽癌中NFκB是否由p65同源二聚体组成,并由此分析LMP1对p65表达水平的影响,进而深入研究调控p65表达及活化的机制,从而为阐述EB病毒致鼻咽癌的可能机理并提出鼻咽癌“靶分子”治疗新策略提供重要的实验依据和理论支持。
基金项目:国家自然科学基金(39700170);国家自然科学基金重点项目(39830410);国家重点基础研究发展规划“恶性肿瘤发生发展的基础研究”(G1998051201)及美国中华医学基金资助(CMB96655)
作者简介:廖伟(1969-),女,湖南娄底人,湖南医科大学肿瘤研究所助理研究员,博士研究生,研究鼻咽癌癌变的分子机理。
, 百拇医药
联系人:曹亚,Tel:0731-4474411-2361;Fax:0731-4470589;Email:ycao98@public. cs.hn.cn
参考文献:
[1]Ranjan S, David Baltimore. Multiple nuclear factors interact with the immunoglobulin enhancers[J]. Cell, 1986, 46: 705-716.
[2]Sankar G, Michael J, Elizabeth B K. NFκB and Rel protein: Evolutionarily conserved mediators of immune responses[J]. Annu Rev Immunol, 1998, 16: 225-260.
, 百拇医药
[3]Ulrich S, Guido F, Keith B. Structrue, regulation and function of NFκB[J]. Annu Rev Cell Biol, 1994, 10: 405-455.
[4]Eliopoulos A G, et al. Epstein-Barr virus-encoded LMP1 and CD40 mediate IL-6 production in epithelial cells via an NF-kappa B pathway involving TNF receptor-associated factors[J]. Oncogene, 1997, 14(24): 2899-2916.
[5]Izumi K M, et al. The Epstein-Barr virus oncogene product, latent membrane protein 1, engages the tumor necrosis factor receptor-associated death domain protein to mediate B lymphocyte growth transformation and activate NF-kappa B[J]. Proc Natl Acad Sci USA, 1997, 94: 1447-1452.
, http://www.100md.com
[6]祝和成,姚开泰,李桂源,等.4株鼻咽癌上皮细胞株的建立及其生物学特性[J].湖南医科大学学报,1992,17(2):103-107.
[7]廖伟,易红,李晓艳,等.Tet调控的LMP1导入鼻咽癌细胞的研究[J].生物物理与生物化学学报,1999,(3):95-98.
[8]Isao K, Toshiya S, James B, et al. Ablation of transplanted HTLV-1 Tax-transformed tumors in mice by antisense inhibition of NFκB[J]. Science, 1992, 258: 1792-1795.
[9]Ichikawa K, Deroot I J. Defetoff S, et al. Nuclear thyriod hormone receptors in cultured human fibroblasts: improved method of isolation, partial characterization, and interaction with chromatin[J]. Metabolism, 1986, 35(9): 861-864.
[10]Fields B N, et al. Fields Virology[M]. New York: Raven Press, 1996, 2343-2396.
收稿日期:1999-08-12;修回日期:1999-12-21, 百拇医药
单位:湖南医科大学肿瘤研究所,湖南 长沙 410078
关键词:潜伏膜蛋白1(LMP1);核转录因子κB;鼻咽癌
病毒学报000302 摘要:为了探讨EB病毒潜伏膜蛋白1(LMP1)的致瘤机制,对鼻咽癌细胞中LMP1通过核转录因子κB(NFκB)介导的信号传导途径在鼻咽癌变中的意义进行了研究。利用LMP1受四环素衍生物强力霉素诱导表达的鼻咽癌细胞Tet-on-LMP1 HNE2,通过NFκB报道基因分析法、凝胶迁移率分析(EMSA)及细胞集落形成率等方法,结合硫代磷酸反义寡核苷酸阻断技术,证实LMP1增强鼻咽癌细胞NFκB的DNA结合活性和反式激活活性,这种增强的活性可被LMP1及NFκB p65硫代磷酸反义寡核苷酸阻断,而NFκB p50仅阻断DNA结合活性,对反式激活活性无影响。同时,LMP1及NFκB p65硫代磷酸反义寡核苷酸可部分逆转鼻咽癌细胞的恶性表型,而反义p50这种效应不显著。这些结果表明,LMP1在鼻咽癌中通过NFκB信号传导途径可能参与了鼻咽癌变,NFκB p65可能是主要的效应者。
, 百拇医药
中图分类号:R373 文献标识码:A
文章编号:1000-8721(2000)03-0198-05
The Epstein-Barr Virus Encoded Latent Membrane Protein 1 Promotes Nuclear Factor-kappa B Activity in a Nasopharyngeal Carcinoma Cell Line
LIAO Wei,TANG Min,DENG Xi-yun,CAO Ya
(Cancer Research Institute, Hunan Medical University, Changsha, Hunan 410078, China)
Abstract: As an oncogenic protein, the Epstein-Barr virus encoded latent membrane protein 1(LMP1)may play a key role in nasopharyngeal carcinogenesis through its signal transduction pathway, especially NF-κB pathway. In order to attest this hypothesis, a dual-stable LMP1 integrated nasopharyngeal carcinoma cell line with Tet-on regulating system designated Tet-on-LMP1 HNE2, in which expression of LMP1 could be dose-dependent regulated by doxycycline, was used. It was found that NFκB activity was 3-10 folds high at LMP1 induced statement by reporter gene analysis, and this result was further confirmed by electrophoresis mobility shift assay which showed NFκB accumulated in nuclear when LMP1 expression was induced. When Tet-on-LMP1 HNE2 cells were co-incubated with phosphorothioate oligonucleotides (PS-ODNs) to antisense LMP1, NFκB p65 and NFκB p50 at various times, NFκB activity curves showed that antisense LMP1 blocked NFκB activity at earlier time than antisense p65 did, and p50 showed no block effect. What's more, PS-ODNs to antisense LMP1 and NFκB p65, but not p50, partially reversed malignant phenotype of Tet-on-LMP1 HNE2 boosted by LMP1 expression. All of these deduced that LMP1 could participate in the nasopharyngeal carcinogenesis by improving NFκB activity in NPC, and NFκB, of which p65 in particular, has the potency used as a molecular target to NPC gene therapy.
, 百拇医药
Key words: Epstein-Barr virus latent membrane protein 1; nuclear factor-kappa B(NFκB); nasopharyngeal carcinoma
核转录因子κB(NFκB)是近年来研究的热点,因与免疫球蛋白kappa轻链基因(Igκ基因)的3′末端增强子结合而得名[1]。NFκB属于一个高度保守的反式激活因子家族,由多个成员组成,包括Dif、Dorsal、Relish、v-rel、c-rel、p50(NF-κB1)、p65(RelA)、RelB和p52。家族中每个成员都包含N末端由300个氨基酸组成的保守序列,称rel同源区域(rel homology domain,RHD)。该区域负责DNA-binding、二聚体聚合及同IκB家族结合等功能,其中Dif、Dorsal、Relish、v-rel、c-rel、p65(RelA)和RelB C末端具有与反式激活相关的部位(transactivation site),p52与p50则没有。NFκB家族成员之间可形成同源二聚体如p50∶p50,或异源二聚体如p50∶p65等。在大多数细胞类型中,NFκB表现为p50∶p65二聚体形式,其他类型的二聚体同时也有低水平的表达[2]。
, 百拇医药
位于胞浆内的NFκB一般与NFκB活性抑制因子IκBα结合,其Rel同源区域被遮蔽,从而不能进入核内发挥反式激活活性。在外来刺激因素,如UV紫外线、氧自由基、病毒和细菌内毒素等的作用下,IκBα被磷酸化并降解,NFκB与IκBα解离,由胞浆转位入细胞核,启动具有κB结合位点的基因转录表达。作为重要的转录调控因子,NFκB调节很多的基因表达,这些基因与细胞增殖、分化、免疫反应、凋亡、转化有着密切关系[3]。
EB病毒是已被认为与鼻咽癌发病密切相关的因素,其编码的潜伏膜蛋白LMP1由于具有潜在的致瘤作用,而成为近年来EB病毒致瘤机制研究的热点。研究发现,LMP1可能类似于肿瘤坏死因子家族成员CD40,在细胞增殖及凋亡的信号传导途径中具有重要的作用。在LMP1所介导的细胞调控网络系统中,NFκB处于信号传导枢纽。近年来研究发现,LMP1蛋白至少可以通过两条途径磷酸化IκBα并活化NFκB:一条为肿瘤坏死因子相关蛋白(tumor necrosis factor receptor associated factors,TRAFs)途径[4];另一条为肿瘤坏死因子受体死亡结构域(tumor necrosis factor receptor associated death domain protein,TRADD)途径[5]。其中TRAFs与CTAR1结合,激活大约25%的NFκB活性;TRADD与CTAR2结合,75%的NFκB的活化与此相关。当LMP1的6个跨膜结构域聚合后,TRAFs和TRADD与LMP1上各自的结构域结合,激活Iκ激酶,Iκ激酶活化后,促使IκBα磷酸化,活化NFκB。
, 百拇医药
作为与鼻咽癌发病密切相关的EB病毒瘤基因,LMP1能通过磷酸化IκBα而活化NFκB,从而提示我们探讨在鼻咽癌中LMP1能否调节NFκB的活性及LMP1是否通过NFκB信号传导途径参与鼻咽癌变的过程等问题,对于阐述EB病毒致鼻咽癌的可能机理并提出鼻咽癌“靶分子”治疗新策略具有较为重要的意义。
材料与方法
1 细胞系 鼻咽癌细胞系HNE2是湖南医科大学肿瘤研究所建立的EB病毒阴性的低分化鳞癌细胞株[6]。PCR进一步证实其中LMP1为阴性。经两轮稳定的转染,将四环素基因调控系统导入其中,构建LMP1受四环素衍生物强力霉素诱导表达的鼻咽癌细胞系pTet-on-LMP1 HNE2[7]。细胞培养液为含15%小牛血清的RPMI-1640。37℃ 5%CO2维持200mg/L G418和50mg/L潮霉素以筛选药物浓度。
2 硫代磷酸化寡脱氧核苷酸有义和反义NFκB p65及p50的设计 硫代磷酸化修饰的寡脱氧核苷酸是进行反义核酸干预的有效方法。根据文献[8],设计针对p65及p50的3′末端硫代磷酸化修饰的有义和反义寡脱氧核苷酸,NFκB p65有义寡脱氧核苷酸:5′-CGGCCATGGACGAACTGTTC-3′,反义寡脱氧核苷酸:5′-GAACAGTTCGTCCACATGGCCCA-CTTC-3';NFκBp50有义寡脱氧核苷酸;5'-ATCTTCACCATGGCAGCAGA-3',反义寡脱氧核苷酸:5'-ATCGTCTGCCATGGTGAAGAT-3'。以上寡脱氧核苷酸由美国国立癌症研究院DNA重组实验室合成,使用浓度为20mg/L浓度。
, 百拇医药
3 NFκB报道基因分析 用NFκB报道基因质粒pNFκB-luc的荧光素酶表达分析NFκB的活性。pNFκB-luc包含螟蛾科荧光素酶基因,它的表达受CMV启动子调控,在CMV启动子中插入了来源于HIV LTR的κB元件。用LipofecTAMINE的方法将pNFκB-luc一过性转染入细胞,转染12~48h后,按荧光素酶分析系统的说明,加入100μl 1×裂解缓冲液,室温下裂解15min,用细胞刮子将细胞刮下,吸至离心管内,12?000r/min离心,取20μl上清液,加入100μl反应底物(Luciferaly-CoA),迅速推入液闪仪(BECKMAN LS5000 TA)单光子档,检测cpm值。间隔10s,每一个样本读两个数值,取均值。
4 凝胶迁移率分析(electrophoresis mobility shift assay,EMSA) 细胞核蛋白的提取参照Ichikawa[9]等的方法进行,收集细胞,冷PBS洗1次,约107个细胞用1ml SMTD-PMSF(0.25mol/L蔗糖,1mmol/L MgCl2,20mmol/L Tris-HCl pH7.85,1mmol/L DTT,0.1mmol/LPMSF)重悬细胞,玻璃匀浆器匀浆4次,加入Triton X-100至终浓度1%,再匀浆2次,离心(1?000g×10min),弃上清,1ml含0.5%Triton X-100的SMTD-PMSF重悬沉淀,玻璃匀浆器再匀浆2次,离心,沉淀再用1ml含1mmol/L CaCl2的SMTD-PMSF洗1次,离心,用与细胞核等体积的KMTD(0.3%KCl,1mmol/L,MgCl2,10mmol/L Tris-HCl pH8.0,1mmol/L DTT)室温温育1h,每隔15min剧烈振动1次,15?000g×15min离心,保留上清核提取物。蛋白浓度用BCA检测试剂(Pierce Chemical Co,Rockford,IL)测定。EMSA分析按照Promega公司Gel shift assay kit操作手册的说明进行。首先用γ-32P-ATP在T4多核苷酸激酶的作用下,连接至NFκB寡核苷酸,37℃ 20min EDTA终止反应,加入89μl TE缓冲液,用G-25柱层析方法纯化探针。约5μg核蛋白与Kit中的结合缓冲液(binding buffer)作用10min,同时以Kit中提供的HeLa细胞核蛋白10μg作为阳性对照;加入1μl纯化后的32P标记NFκB consensus oligo,室温放置约3h,加入1μl加样缓冲液,于4%聚丙烯酰胺凝胶(acrylamide∶bis为39∶1)进行电泳分离。待加样缓冲液中的染料溴酚蓝距电泳槽底部1/4距离时,终止电泳。用带有增感屏的暗闸进行X光片显影、曝光。
, 百拇医药
5 细胞集落形成分析 配制0.5%的琼脂,以该琼脂为底层,将dox(+)、dox(-)的Tet-on-LMP1 HNE2、以20μg/ml正义或反义LMP1及p65、p50处理24h后的Tet-on-LMP1 HNE2 2×104个细胞加入培养基,再用此培养基稀释0.5%的琼脂至0.33%,作为上层琼脂。每组种3个平皿,置5%CO2 37℃培育10天,显微镜下用随机取样法观察每一平皿5个1cm2内克隆数目(8个细胞以上为一集落)及大小。
结 果
1 LMP1促进NFκB的反式激活活性将Tet-on-LMP1 HNE2细胞在dox(-)、dox(+)及分别加入硫代磷酸化修饰的反义或正义NFκB p65的状态下培养36h,用NFκB报道质粒检测NFκB的反式激活活性。由图1可看到,在dox(+)状态或dox(+)状态加上正义NFκB p65,NFκB具有较高的反式激活活性;dox(-)或dox(+)加上反义NFκB p65,则NFκB反式激活活性下降;作为对照,未转染Tet-on调控系统的HNE2即使在dox(+)状态,NFκB反式激活活性也较低。这说明LMP1可促进NFκB的反式激活活性,这种作用可被反义NFκB p65所阻断。
, 百拇医药
图1 LMP1增强NFκB的反式激活活性
Figure 1 LMP1 increased transactivational activity of NFκB
Tet-on-LMP1 HNE2 cell with or without dox, or coincubated with sense and antisense NFκB p65 for 36h was analyzed by means of NFκB reporter gene to detect NFκB transactivational activity. This activity was higher in cells with dox(+), in which expression of LMP1 was induced, and could be blocked by antisense NFκB p65.
, 百拇医药 2 LMP1对NFκB活性调节的时间效应
在证实LMP1促进NFκB反式激活活性的基础上,为了进一步阐明LMP1对NFκB反式激活活性调节的时间效应,并观察与p65和p50对NFκB反式激活活性的影响的差别,以PS-ODNs反义LMP1,p65和p50 20μg/ml与诱导和非诱导状态的Tet-on-LMP1 HNE2细胞共温育,并检测温育后12h、24h、36h和48hNFκB活性,可以看到,反义LMP1导入后,在所有的时间点NFκB活性都较低(图2A);反义p65 PS-ODNs则在导入后36h才产生抑制效应,至48h活性有所回升(图2B)。而反义p50 PS-ODNs在48h内对NFκB的活性几乎都不产生影响(图2C)。
图2 LMP1对NFκB反式激活活性调节的时间效应
Figure 2 Time-course of LMP1 mediated NFκB transactivational activity
, 百拇医药
(A)Antisense LMP1 blocked the NFκB activity. 2μg/ml LMP1 antisense plasmid was transiently transfected into pTet-on-LMP1 HNE2 cell line with or without 2μg/ml dox. Markedly blocked effect was shown at least 12h after pNFκB-luc transfection. (B)Antisense P65 oligonucleotide blocked the NFκB activity. 20μg/ml P65 antisense oligonucleotide was added to the medium of pTet-on-LMP1 cells with or without 2μg/ml dox at indicated time. The most distinct blocked effect was shown at 36h after pNFκB-luc transfection. However, p50 antisense oligonucleotide didn't block the NFκB transactivational activity(C).
, http://www.100md.com
3 LMP1、NFκB p65和NFκB p50调节NFκB的DNA结合活性
LMP1、NFκB p65可以调控NFκB的反式激活活性,而NFκB p50则无这种作用。而这3个因子对NFκB的DNA结合活性是否有影响,利用凝胶移动迁移率(EMSA)进行了进一步探讨。EMSA的结果显示,HeLa阳性对照呈现明显的NFκB两条信号,而对于Tet-on-LMP1 HNE2细胞,在dox(+)状态下,NFκB的DNA结合活性增强,加入硫代磷酸化修饰的反义(antisense)LMP1,p65和p50共温育36h后,这种结合活性显著下降(图3)。
4 LMP1和NFκB p65和NFκB p50对鼻咽癌恶性表型的影响
倒置显微镜下观察dox(+)、dox(-)的Tet-on-LMP1 HNE2,dox(+)加上20μg/ml正义或反义LMP1和dox(+)加p65、p50处理36h后的Tet-on-LMP1 HNE2的细胞集落形成情况,可以看到,在dox(+)状态下细胞集落最为密集,且集落体积较大;在dox(-)、反义LMP1及p65处理后,则细胞集落明显稀疏,体积较小,而反义p50则作用相对不明显(图4)。
, 百拇医药
图3 LMP1对NFκB结合活性的调节
Figure 3 NFκB binding activity regulated by LMP1
NFκB binding activity blocked by antisense LMP1, antisense p65 and antisense p50. NFκB binding activity in Tet-on-LMP1 HNE2 cell line following dox, antisense LMP1, antisense p65 and antisense p50 treatment was analyzed by means of EMSA. Tet-on-LMP1 HNE2 cell line was plated in ten 100-mm cell culture dishes(1×106 cells/dish)in 1640 with 10% fetal bovine serum(FBS)and cultured for 4h. The cell medium was replaced by 1640 with 10% FBS and with dox, antisense LMP1, antisense p65 and antisense p50 and cultured for 36h. 10μg of nuclear protein purified from cells were incubated with 3×104 cpm 32P-labeled consensus NFκB oligonucleotide and separated by 5% acrylamide gel electrophoresis as described in “Materials and Methods”.
, 百拇医药
图4 细胞集落形成检测LMP1、p65和p50对Tet-on-LMP1 HNE2鼻咽癌细胞系恶性表型的影响
Figure 4 Colony forming of Tet-on-LMP1 HNE2 cell line treated as conditions indicated
A. dox(+); B. dox(-); C. dox(+)with PS-ODN LMP1 treated; D. dox(+)with p65 PS-ODN LMP1 treated; E. dox(+)with PS-ODN p50 treated. Experimental procedure was described in detail in "Materials and Methods".
讨 论
LMP1的致瘤机制近年来一直是一个研究的热点。Kieff[10]于1996年提出,LMP1可能类似于肿瘤坏死因子受体家族成员CD40,通过参与细胞内信号传导网络,而参与细胞增殖或凋亡的调控。目前已知LMP1至少与3条信号传导通路有关:①通过TRAFs/TRADD活化NFκB;②通过SEK-JNK1及Ras-Rafl-MER-Erk1/2活化AP-1;③通过JAK-STAT途径调节相关基因的表达。其中LMP1通过NFκB调控其靶基因的表达尤为重要。
, 百拇医药
NFκB的活化可能与许多肿瘤的发生有关。将v-Rel置于T细胞特异性启动子的控制下产生的转基因小鼠会发生T细胞淋巴瘤。此外,NFκB可分别与HTLV-1逆反式激活病毒和HBV病毒中的Tax及x基因产物结合,在它们的致瘤中产生作用。在人T细胞和B细胞肿瘤中大约有1%~10%的Rel或IκBα家族基因发生改变。人乳腺癌和多发性骨髓瘤中均可见NFκB的点突变活化[3]。
本工作利用一株LMP1受四环素衍生物强力霉素诱导表达的鼻咽癌细胞株Tet-on-LMP1 HNE2,探讨了LMP1在鼻咽癌细胞中调控NFκB的活性,并通过这种活性的调控,对鼻咽癌细胞恶性表型产生的影响。
NFκB的活性包括DNA结合活性和反式激活活性两种。用NFκB报道基因分析法可检测其反式激活活性,而凝胶迁移率分析显示的为其DNA结合活性。研究显示,尽管LMP1、p65和p50的反义核苷酸都降低NFκB的DNA结合活性,但三者对其反式激活活性的影响是不同的。LMP1可促进NFκB的反式激活活性,这种作用被反义NFκB p65所阻断;反义LMP1导入后,NFκB的活性在所有的时间点都较低;反义p65 PS-ODNs则在导入后36h才产生抑制效应,而48h活性有所回升。而反义p50 PS-ODNs在48h内对NFκB的反式激活活性几乎都不产生影响。这说明LMP1通过磷酸化IκBα,对已存在于胞浆内的NFκB的活化较早期发生,而反义p65对NFκB活性的影响,可能是先通过抑制p65 RNA合成,再降低p65蛋白的翻译,最终影响NFκB的活性,反义p50的干预对NFκB活性不产生作用,进一步提示p50对NFκB的反式激活活性无影响。
, http://www.100md.com
细胞集落形成实验证实LMP1可促进细胞集落形成率,而反义LMP1和反义p65可部分阻断这种恶性表型,反义p50则作用不显著。
由此我们推断,EB病毒LMP1通过活化NFκB,继而调节相关基因的表达,在鼻咽癌的癌变过程中可能发挥了重要作用。其中,反义p65寡核苷酸对NFκB反式激活活性的阻断及对细胞集落形成的影响,提示应进一步探讨鼻咽癌中NFκB是否由p65同源二聚体组成,并由此分析LMP1对p65表达水平的影响,进而深入研究调控p65表达及活化的机制,从而为阐述EB病毒致鼻咽癌的可能机理并提出鼻咽癌“靶分子”治疗新策略提供重要的实验依据和理论支持。
基金项目:国家自然科学基金(39700170);国家自然科学基金重点项目(39830410);国家重点基础研究发展规划“恶性肿瘤发生发展的基础研究”(G1998051201)及美国中华医学基金资助(CMB96655)
作者简介:廖伟(1969-),女,湖南娄底人,湖南医科大学肿瘤研究所助理研究员,博士研究生,研究鼻咽癌癌变的分子机理。
, 百拇医药
联系人:曹亚,Tel:0731-4474411-2361;Fax:0731-4470589;Email:ycao98@public. cs.hn.cn
参考文献:
[1]Ranjan S, David Baltimore. Multiple nuclear factors interact with the immunoglobulin enhancers[J]. Cell, 1986, 46: 705-716.
[2]Sankar G, Michael J, Elizabeth B K. NFκB and Rel protein: Evolutionarily conserved mediators of immune responses[J]. Annu Rev Immunol, 1998, 16: 225-260.
, 百拇医药
[3]Ulrich S, Guido F, Keith B. Structrue, regulation and function of NFκB[J]. Annu Rev Cell Biol, 1994, 10: 405-455.
[4]Eliopoulos A G, et al. Epstein-Barr virus-encoded LMP1 and CD40 mediate IL-6 production in epithelial cells via an NF-kappa B pathway involving TNF receptor-associated factors[J]. Oncogene, 1997, 14(24): 2899-2916.
[5]Izumi K M, et al. The Epstein-Barr virus oncogene product, latent membrane protein 1, engages the tumor necrosis factor receptor-associated death domain protein to mediate B lymphocyte growth transformation and activate NF-kappa B[J]. Proc Natl Acad Sci USA, 1997, 94: 1447-1452.
, http://www.100md.com
[6]祝和成,姚开泰,李桂源,等.4株鼻咽癌上皮细胞株的建立及其生物学特性[J].湖南医科大学学报,1992,17(2):103-107.
[7]廖伟,易红,李晓艳,等.Tet调控的LMP1导入鼻咽癌细胞的研究[J].生物物理与生物化学学报,1999,(3):95-98.
[8]Isao K, Toshiya S, James B, et al. Ablation of transplanted HTLV-1 Tax-transformed tumors in mice by antisense inhibition of NFκB[J]. Science, 1992, 258: 1792-1795.
[9]Ichikawa K, Deroot I J. Defetoff S, et al. Nuclear thyriod hormone receptors in cultured human fibroblasts: improved method of isolation, partial characterization, and interaction with chromatin[J]. Metabolism, 1986, 35(9): 861-864.
[10]Fields B N, et al. Fields Virology[M]. New York: Raven Press, 1996, 2343-2396.
收稿日期:1999-08-12;修回日期:1999-12-21, 百拇医药