Β―ι–‘“ΰΊΚ”’ΒΦ–Γ σ…ζΨΪœΗΑϊΒρΆωΒΡ―–ΨΩ*
Ής’ΏΘΚ–λΫΓ –μ‘ω¬Μ ΫΣ”ΔΧΈ «°œΰίΦ ΜΤ”ώήΏ
ΒΞΈΜΘΚ–λΫΓ –μ‘ω¬Μ ΫΣ”ΔΧΈ «°œΰίΦ ΜΤ”ώήΏ(÷–Ιζ“Ϋ―ßΩΤ―ß‘Κ ÷–Ιζ–≠ΚΆ“ΫΩΤ¥σ―ßΜυ¥Γ“Ϋ―ß―–ΨΩΥυΘ§±±Ψ© 100005)
ΙΊΦϋ¥ ΘΚ…ζΨΪœΗΑϊΒρΆωΘΜBcl-2ΒΑΑΉΘΜBaxΒΑΑΉ
ΫβΤ ―ß±®990121
ΓΨ’Σ“ΣΓΩ ΡΩΒΡ ―–ΨΩ ÷ θ“ΰΊΚΥυ÷¬≥…Ρξ–Γ σ…ζ÷≥œΗΑϊΒρΆωΦΑœύΙΊΒςΫΎ“ρΉ”bcl-2ΓΔbaxΒΑΑΉ‘Ύ«ζœΗΨΪΙή÷–ΒΡΕ®ΈΜΓΔ±δΜ·ΓΘ ΖΫΖ® “‘Ρ©ΕΥΆ――θΚΥΧ«ΚΥΥαΉΣ“ΤΟΗΫιΒΦΒΡdUTP»±ΩΎΡ©ΕΥ±ξΦ«Ζ®(terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling method,TUNEL)Φλ≤βΒρΆωΒΡ…ζ÷≥œΗΑϊΘ§…ζΈοΥΊ-ΩΙ…ζΈοΥΊΒΑΑΉDCS(biotin-avidin D cell sorter)ΧεœΒΦδΫ”Οβ“Ώ”ΪΙβΖ®Φλ≤βBcl-2ΓΔBaxΒΑΑΉ‘Ύ«ζœΗΨΪΙή÷–ΒΡΕ®ΈΜΓΔΖ÷≤ΦΓΘ ΫαΙϊ “ΰΊΚ θΚσ3dΘ§ ÷ θ≤ύΊΚΆη÷ΊΝΩΦΑœΗΑϊΒρΆω ΐΝΩ”κΕ‘≤ύΈόΟςœ‘«χ±πΓΘΕχ6ΓΪ15dΘ§ΊΚΆη÷ΊΝΩΟςœ‘Φθ«αΘ§ΒρΆωœΗΑϊΟςœ‘‘ωΕύΘ§“‘ΨΪΡΗœΗΑϊΓΔΨΪΉ”œΗΑϊΈΣ÷ςΓΘ‘Ύ’ΐ≥Θ–Γ σ«ζœΗΨΪΙή÷–Θ§bcl-2¥σΝΩ±μ¥οΘ§≥ …Δ‘ΎΒΡΩ≈ΝΘΉ¥¥φ‘Ύ”ΎΗςΦΕ…ζΨΪœΗΑϊ÷–Θ§“‘ΨΪΉ”œΗΑϊΑϊ÷ ΈΣ÷ςΓΘBaxΒΑΑΉ±μ¥οΝΩΫœ…ΌΘ§“Μ≤ΩΖ÷“≤“‘…Δ‘ΎΒΡΩ≈ΝΘ¥φ‘Ύ”ΎΗςΦΕ…ζΨΪœΗΑϊ÷ ÷–Θ§Νμ“Μ≤ΩΖ÷‘ρΫœ÷¬ΟήΒΊΖ÷≤Φ”Ύ«ζœΗΨΪΙήΜυΒΉ≤ΩΓΘ“ΰΊΚ θΚσ3dΘ§Bcl-2ΓΔBaxΒΡ±μ¥ο”κΕ‘≤ύΈόΟςœ‘«χ±πΘΜ6ΓΪ15dΘ§Bcl-2Οςœ‘œ¬ΫΒΘ§BaxΟςœ‘‘ωΦ”ΓΘ Ϋα¬έ “ΰΊΚΟςœ‘”’ΒΦ…ζ÷≥œΗΑϊΒρΆωΘ§Bcl-2ΓΔBaxΒΑΑΉΕ‘…ζ÷≥œΗΑϊΒΡΒρΆω”–÷Ί“ΣΒςΫΎΉς”ΟΘ§Χα ΨΈ¬Ε»ΓΔBcl-2ΓΔBaxΒΑΑΉ‘ΎΨΪΉ”ΖΔ…ζΙΐ≥Χ÷–Ε‘…ζΨΪœΗΑϊΒΡ÷ςΕ·ΆΥΜ·Ω…Ρή”–÷Ί“ΣΉς”ΟΓΘ
, ΑΌΡ¥“Ϋ“©
CRYPTORCHIDISM INDUCES TESTICULAR GERM CELL APOPTOSIS
IN MICE TESTES AND CHANGES OF Bcl-2,Bax PROTEIN EXPRESSION
Xu Jian, Xu ZengluΓς, Jiang Yingtao, Qian Xiaojing, Huang Yuling
(Institute of Basic Medical Sciences, CAMS and PUMC, Beijing)
ΓΨAbstractΓΩ Objective To investigate the effect of experimentally induced cryptorchidism on apoptosis in testes of adult mice and the distribution changes of Bcl-2, Bax protein. Methods Total DNA was isolated from testes and was electrophoretically separated in 1.5% agarose gel, TUNEL method, biotin-avidin DCS system indirect immunofluorescence techniques were used. Results DNA fragmentation, increasing apoptotic germ cells, in coordination with the loss of testicular weight, were observed on 6-15d in the cryptorchid testes. By TUNEL staining it was noted that postive cells were most frequently among primary spermatocytes and round spermatids. Bcl-2, Bax protein mainly distributed in cytoplasm of germ cells. With the increase of apoptosis, Bcl-2 was down and Bax was up regulated. Conclusion Temperature, Bcl-2, and Bax protein play an important role in male germ cell apoptosis.
, ΑΌΡ¥“Ϋ“©
ΓΨKey wordsΓΩ Apoptotic germ cells; Bcl-2 protein; Bax protein
’ΐ≥ΘΒΡ…ζΨΪΙΐ≥Χ÷–Θ§¥φ‘ΎΉ≈…ζ÷≥œΗΑϊΒΡ÷ςΕ·ΆΥΜ·œ÷œσΘ§≥ΛΤΎ“‘ά¥ ήΒΫ―–ΨΩ’ΏΒΡ÷Ί ”Θ§ΤδΜζ÷Τ»‘≤Μ«ε≥ΰΓΘΫϋΦΗΡξά¥Θ§ΙζΆβ“Μ–©―–ΨΩ’ΏΩΣ ΦΫΪ’β÷÷÷ςΕ·ΆΥΜ·”κœΗΑϊΒρΆωΝΣœΒ‘Ύ“ΜΤπΘ§ΚήΕύ Β―ι“≤±μΟςΒρΆωΩ…Ρή «…ζ÷≥œΗΑϊ÷ςΕ·ΆΥΜ·ΒΡ“Μ÷÷Μζ÷Τ[1Θ§2]ΓΘ”…”Ύœ¬«πΡ‘-¥ΙΧε-–‘œΌ÷αΕ‘…ζ÷≥ΒΡΟςœ‘ΩΊ÷ΤΉς”ΟΘ§¥σΕύ ΐ Β―ιΦ·÷–”Ύ”–ΙΊΡΎΖ÷ΟΎΕ‘…ζ÷≥œΗΑϊΒρΆωΒςΫΎΖΫΟφΒΡ―–ΨΩΘ§ΕχΊΚΆηΜΙ±Ί–κ‘ΎΒΆ”ΎΧεΈ¬4ΓΪ5ΓφΒΡ“θΡ“÷–≤≈Ρή’ΐ≥ΘΖΔ”ΐΘ§Έ¬Ε»“≤ «÷Ί“ΣΒΡΒςΫΎ“ρΥΊ÷°“ΜΓΘ“ρ¥ΥΘ§Έ“Ο«“‘ΒΞ≤ύ ÷ θ“ΰΊΚΗΡ±δΨΪΉ”ΖΔ…ζΥυ–η“ΣΒΡ’ΐ≥ΘΈ¬Ε»ΒΡΖΫΖ®ά¥Ϋ®ΝΔ…ζ÷≥œΗΑϊΒρΆωΒΡΡΘ–ΆΓΘ
–μΕύ―–ΨΩ±μΟςbcl-2Μυ“ρΦ“Ήε «ΒςΫΎœΗΑϊΒρΆωΒΡ“ΜΗω÷Ί“ΣΜυ“ρΦ“ΉεΘ§ΒΪΦ“Ήε≥…‘±Ε‘ΒρΆωΒΡΉς”Ο»¥”–Οςœ‘≤ΜΆ§ΓΘ“Μάύ¥ΌΫχœΗΑϊΒρΆω»γbaxΓΔbcl-xsΒ»Θ§Νμ“Μάύ‘ρ“÷÷ΤΒρΆω»γbcl-2ΓΔbcl-xΒ»[3]ΓΘbcl-2Μυ“ρΦ“ΉεΕ‘÷ΉΝωœΗΑϊΓΔΝήΑΆœΗΑϊΒρΆωΉς”ΟΒΡ―–ΨΩΚήΕύΘ§Ε‘…ζ÷≥œΗΑϊΒρΆωΒΡΉς”ΟΘ§ΉνΫϋ“≤”–Ηω±π±®Βά[4]ΓΘΈ“Ο«‘ρ―–ΨΩΝΥBcl-2ΓΔBaxΒΑΑΉ‘Ύ’ΐ≥Θ–Γ σΊΚΆη«ζœΗΨΪΙή÷–ΒΡΕ®ΈΜΦΑΖ÷≤ΦΘ§ΦΑΤδ‘Ύ…ζΨΪœΗΑϊΒρΆω ±Θ§±μ¥οΒΡΗΡ±δΓΘ
, http://www.100md.com
≤ΡΝœΚΆΖΫΖ®
1.Ε·Έοά¥‘¥ΦΑΕ·ΈοΡΘ–ΆΒΡΫ®ΝΔ
÷–Ιζ“Ϋ―ßΩΤ―ß‘ΚΕ·ΈοΥυΧαΙ©ΒΡ8ΓΪ10÷ή(26ΓΪ30g)ΫΓΩΒ–έ–‘άΞΟς–Γ σΓΘΈλΑΆ±»ΆΉΗΙ«ΜΉΔ…δ¬ιΉμ(30mg/kg)Θ§ΖλΚœΒΞ≤ύΗΙΙ…ΙΒΙήΘ§ Ι–Γ σΊΚΆη≤ΜΡή¥”ΗΙ«Μœ¬ΫΒ÷Ν“θΡ“Θ§Έ¥ ÷ θ≤ύΦΑ’ΐ≥Θ–Γ σΒΡΊΚΆηΉςΕ‘’’Θ§ θΚσ3ΓΔ6ΓΔ8ΓΔ10ΓΔ12ΓΔ15d»Γ≤ΡΘ§ΟΩΉιΕ·Έο6ΓΪ8÷ΜΓΘ
2. ‘ΦΝ
œΗΑϊΒρΆω ‘ΦΝΚ–(ClontechΙΪΥΨ)Θ§ΆΟΩΙ–Γ σBcl-2ΓΔBaxΕύΩΙ(Santa Cruz BiotechnologyΙΪΥΨΘ§ΙΚ”Ύ±±Ψ©÷–…Ϋ…ζΈοΦΦ θ”–œόΙΪΥΨ)Θ§…ζΈοΥΊΜ·ΒΡ…Ϋ―ρΩΙΆΟIgGΘ§”ΪΙβΥΊ-ΩΙ…ζΈοΥΊΒΑΑΉDCSΓΔ…Ϋ―ρ―Σ«ε(Vector Laboratories, Burlingame),ppd(phenylenediamine)ΓΔagaroseΓΔRNase AΓΔprotein kinase (SigmaΙΪΥΨ)ΓΘ
, http://www.100md.com
3.ΊΚΆη≥Τ÷ΊΦΑHE»Ψ…Ϊ
–Γ σΨ±ΉΒΆ―Ψ Ζ®¥ΠΥάΘ§»ΓΥΪ≤ύΊΚΆηΘ§≥Τ÷ΊΓΘ10%÷––‘ΗΘΕϊ¬μΝ÷ΙΧΕ®24hΘ§≥ΘΙφΑϋ¬ώΓΔ«–Τ§ΓΔHE»Ψ…ΪΓΘ
4.Ήι÷·DNAΧα»ΓΦΑ«μ÷§Χ«ΡΐΫΚΒγ”Ψ
¥”¥ΠΥάΒΡ–Γ σ÷–»Γ≥ωΊΚΆη(Ω…-70Γφ±Θ¥φ)Θ§»ΞΑΉΡΛΘ§―–ΥιΘ§Φ”»κ10±ΕΧεΜΐΒΡbuffer(10mmol/L Tris-HCl, pH8.0; 0.1mol/L EDTA,pH8.0;0.5%SDS; 100mmol/L NaCl,4ΓφΙΐ“Ι)Θ§Φ”RNase A÷Ν÷’≈®Ε»40mg/L,37ΓφΘ§1hΘΜΦ”protein kinase÷Ν÷’≈®Ε»100mg/L,50ΓφΘ§3h;10 000gάκ–Ρ30minΘ§…œ«ε”ΟΖ”/¬»Ζ¬≥ιΧα2¥ΈΘ§ΈόΥ°““¥Φ-20ΓφΙΐ“ΙΘ§12 000gάκ–Ρ30minΓΘ’φΩ’≥ιΗ…Θ§TE buffer (10mmol/L Tris HCl pH8.0; 1mmol/L EDTA, pH8.0)»ήΫβΓΘ1.5%«μ÷§Χ«ΡΐΫΚΒγ”ΨΓΘ
, ΑΌΡ¥“Ϋ“©
5.ΒρΆωœΗΑϊΒΡΦλ≤βΦΑΕ®ΝΩ ΐΨί¥Πάμ
Α¥’’ ‘ΦΝΚ–ΥΒΟςΘ§“‘TUNELΖ®Φλ≤βΒρΆωœΗΑϊΘ§œ‘¬Χ…Ϊ”ΪΙβΒΡœΗΑϊΈΣ―τ–‘œΗΑϊΓΘΑ¥Yin[5]Ζ®Ής ΐΨί¥ΠάμΘΚœ‘ΈΔΨΒœ¬»Έ―Γ5ΓΪ10Ηω ”“ΑΘ§ΦΤ ΐ50Ηω«ζœΗΨΪΙήΚα«–ΟφΘ§Κ§ΒρΆωœΗΑϊ’ΏΈΣ―τ–‘«ζœΗΨΪΙήΘ§‘ΌΦΤ ΐ15ΓΪ20Ηω―τ–‘«ζœΗΨΪΙή÷–ΒΡΉήœΗΑϊ ΐΚΆ―τ–‘œΗΑϊ ΐΘ§ΦΤΥψΒρΆωœΗΑϊΒΡΑΌΖ÷±»ΓΘΟΩΗω ±ΦδΒψΦΤΥψ3÷Μ–Γ σΒΡΊΚΆηΘ§ΟΩΗωΊΚΆη ΐ3’≈«–Τ§ΓΘ
6.…ζΈοΥΊ-ΩΙ…ζΈοΥΊΒΑΑΉDCSΦδΫ”Οβ“Ώ”ΪΙβ»Ψ…Ϊ
»Γ–¬œ ΊΚΆηΉι÷·Θ§OCTΑϋ¬ώΘ§“ΚΒΣ÷–άδΕ≥Θ§Κψάδœδ«–Τ§Θ§Κώ10ΠΧmΘ§0.1mol/L PBS≈δ÷ΤΒΡ10%…Ϋ―ρ―Σ«ε4ΓφΖθ”ΐ20minΘ§¬Υ÷ΫΈϋ»ΞΕύ”ύ―Σ«εΘ§Φ”…œ”Ο10%…Ϋ―ρ―Σ«εœΓ ΆΒΡ(1ΓΟ100)ΆΟΩΙ–Γ σBcl-2ΓΔBaxΕύΩΙΘ§4ΓφΙΐ“ΙΘ§“θ–‘Ε‘’’ ûΥΥ≤Ϋ÷ηΘ§άδPBS 4Γφœ¥3¥ΈΘ§ΟΩ¥Έ5minΘ§Φ”…œ10%…Ϋ―ρ―Σ«εœΓ ΆΒΡ(1ΓΟ200)…ζΈοΥΊΜ·…Ϋ―ρΩΙΆΟIgGΘ§4ΓφΖθ”ΐ1hΘ§άδPBS”Ύ4Γφœ¥3¥ΈΘ§ΟΩ¥Έ5minΓΘ“‘”ΪΙβΥΊ-ΩΙ…ζΈοΥΊΒΑΑΉDCS (1ΓΟ200ΧΦΥα«βΡΤΜΚ≥ε“ΚœΓ Ά)4ΓφΖθ”ΐ1hΘ§άδPBS 4Γφœ¥4¥ΈΘ§ΟΩ¥Έ5minΘ§”ΟΚ§10%PBSΚΆ1% ppdΒΡΗ ”ΆΖβΤ§[6]ΓΘ±ξ±Ψ‘ΎOlympusΖ¥…δ Ϋ”ΪΙβœ‘ΈΔΨΒœ¬Ιέ≤λΘ§≤Δ”Ο400 ASA KodakΫΚΨμ…ψΤ§ΓΘ
, http://www.100md.com
Ϋα Ιϊ
1.ΊΚΆη÷ΊΝΩΒΡΗΡ±δ
’ΐ≥Θ σΦΑΕ‘’’≤ύΊΚΆηΒΡ÷ΊΝΩ‘Ύ’ϊΗω Β―ιΙΐ≥Χ÷–ΈόΟςœ‘±δΜ·ΓΘ“ΰΊΚ÷ΊΝΩ‘Ύ θΚσ3d”κΕ‘’’≤ύΈόΟςœ‘±δΜ·ΓΘ θΚσ6dΦθ«α20%Θ§8dΦθ«α29%Θ§10dΦθ«α32%Θ§12dΦθ«α46%Θ§15dΦθ«α31%ΓΘ“ΰΊΚ≤ύ”κΕ‘’’≤ύ÷ΊΝΩΒΡ≤ν±π‘Ύ6ΓΪ15dΟΩΗω ±ΦδΒψΨυ”–œ‘÷χ≤ν“λ(P<0.01)Θ§ΕχΉιΦδΈόœ‘÷χ≤ν“λ(ΆΦ1)ΓΘ
ΆΦ1 “ΰΊΚΦΑΕ‘’’≤ύΊΚΆη÷ΊΝΩΒΡ±δΜ·
Fig.1 Both cryptorchid and control testesweights ( ) are plotted against days of experimental cryptorchidism from 3 to 15.
) are plotted against days of experimental cryptorchidism from 3 to 15.
, http://www.100md.com
2.HE»Ψ…Ϊ
Ε‘’’≤ύΊΚΆη…ζΨΪ…œΤΛΆξ’ϊΘ§ΗςΦΕ…ζΨΪœΗΑϊΦΑΨΪΉ”≈≈Ν–”––ρΓΘ“ΰΊΚ θΚσ3dΈόΟςœ‘±δΜ·ΓΘ θΚσ6ΓΪ15d«ζœΗΨΪΙήΙή«Μ‘ω¥σΘ§…ζΨΪ…œΤΛ±δ±ΓΘ§≤ΜΆξ’ϊΘ§ΨΪΉ”Οςœ‘Φθ…ΌΘ§≤ΔΩ…Φϊ“Μ–©œΗΑϊΒΡ»Ψ…Ϊ÷ Οςœ‘±ΏΦ·Μ·Θ§≥ ΜΖΉ¥ΓΘ“Μ–©«ζœΗΨΪΙή÷–≥ωœ÷ΓΑΕύΚΥΨόœΗΑϊΓ±(multinucleated giant cells)Θ§Φ¥ΕύΗωœΗΑϊΚΥΨέΦ·–Έ≥…“ΜΗω¥σΒΡœΗΑϊΆ≈ΩιΘ§Τδ÷–“Μ–©œΗΑϊΚΥ»Ψ…Ϊ÷ Οςœ‘±ΏΦ·Μ·Θ§≥ ΜΖΉ¥ΓΘΦδ÷ œΗΑϊΚΆ÷ß≥÷œΗΑϊΈό±δΜ·ΓΘ
3.ΡΐΫΚΒγ”Ψ
“ΰΊΚ θΚσ6ΓΔ8ΓΔ10dΩ…ΦϊΓΑDNA ladderΓ±ΆΦœώΘ§œ‘ Ψ”–ΚΥ–ΓΧεΦδDNAΕœΝ―ΒΡ–ΓΤ§ΕΈ¥φ‘ΎΘ§Εχœύ”ΠΒΡΕ‘’’≤ύΓΔ“ΰΊΚ θΚσ3dΦΑ’ΐ≥Θ–Γ σΊΚΆηΟΜ”–Ιέ≤λΒΫ–ΓΤ§ΕΈDNA(ΆΦ2)ΓΘ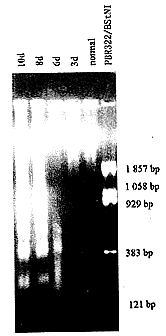
ΆΦ2 DNA«μ÷§Χ«ΡΐΫΚΒγ”ΨΆΦΤΉ
, http://www.100md.com
Fig.2 Agarose gel fractionation of DNA.Low molecular-weight
DNA from both cryptorchid and control testes were fractionated
on 1.5% agarose gel containing 0.5mg/L ethidium bromide.
4.TUNEL»Ψ…Ϊ
’ΐ≥ΘΦΑΕ‘’’≤ύΊΚΆη”–…Ό ΐ«ζœΗΨΪΙή÷–Ω…Ιέ≤λΒΫΗω±πΒρΆωœΗΑϊΘ§÷ς“ΣΈΣ≥θΦΕΨΪΡΗœΗΑϊ(ΆΦ3A)ΓΘ“ΰΊΚ θΚσ3dΈόΟςœ‘±δΜ·Θ§ θΚσ6ΓΪ15d―τ–‘«ζœΗΨΪΙήΦΑ―τ–‘œΗΑϊΨυΟςœ‘‘ωΦ”ΓΘΒρΆωœΗΑϊ÷ς“ΣΑϋά®ΨΪΡΗœΗΑϊΚΆΨΪΉ”œΗΑϊΘ§≤ΩΖ÷ΓΑΕύΚΥΨόœΗΑϊΓ±“≤≥ ―τ–‘ΓΘΈ¥ΦϊΦδ÷ œΗΑϊΦΑ÷ß≥÷œΗΑϊΒρΆω(ΆΦ3B)ΓΘ’ΐ≥ΘΦΑΕ‘’’≤ύΊΚΆη―τ–‘«ζœΗΨΪΙή(Κ§ΒρΆωœΗΑϊΒΡ«ζœΗΨΪΙή)ΈΣ10.7%Θ§―τ–‘œΗΑϊ(ΒρΆωœΗΑϊ)’Φ1.6%ΓΘ“ΰΊΚ θΚσ3d―τ–‘«ζœΗΨΪΙή12%Θ§―τ–‘œΗΑϊ2.1%ΓΘ θΚσ6d―τ–‘«ζœΗΨΪΙή68.6%Θ§―τ–‘œΗΑϊ5.6%Θ§ θΚσ8d―τ–‘«ζœΗΨΪΙή72%Θ§―τ–‘œΗΑϊ5.8%Θ§ θΚσ10d―τ–‘«ζœΗΨΪΙή64.6%Θ§―τ–‘œΗΑϊ4.8%Θ§ θΚσ12d―τ–‘«ζœΗΨΪΙή58.9%Θ§―τ–‘œΗΑϊ4.1%Θ§ θΚσ15d―τ–‘«ζœΗΨΪΙή67.2%Θ§―τ–‘œΗΑϊ3.9%(ΆΦ4AΘ§B)ΓΘ θΚσ3d”κΕ‘’’≤ύΈόΟςœ‘≤ν“λ(P>0.05)ΓΘ θΚσ6ΓΪ15d”κΕ‘’’≤ύΦΑ3dΨυ”–œ‘÷χ≤ν“λ(P<0.05)Θ§6ΓΪ15dΗςΉι÷°ΦδΈόœ‘÷χ≤ν“λ(P>0.05)ΓΘ
, ΑΌΡ¥“Ϋ“©
5.Bcl-2ΓΔBaxΒΑΑΉ‘Ύ«ζœΗΨΪΙή÷–ΒΡΖ÷≤ΦΦΑ±δΜ·
‘Ύ’ΐ≥ΘΦΑΕ‘’’≤ύΊΚΆηΒΡ«ζœΗΨΪΙή÷–Θ§Bcl-2ΒΑΑΉ“‘…Δ‘ΎΒΡΩ≈ΝΘΉ¥¥σΝΩΖ÷≤Φ”ΎΗςΦΕ…ζΨΪœΗΑϊ÷–Θ§“‘ΨΪΉ”œΗΑϊ÷ ΈΣΉνΕύ(ΆΦ3C)Θ§ θΚσ3dΈόΟςœ‘±δΜ·Θ§ θΚσ6ΓΪ15d‘ρΦϊ”ΪΙβΩ≈ΝΘΟςœ‘Φθ…Ό(ΆΦ3D)ΓΘΕχBaxΒΑΑΉ‘Ύ’ΐ≥ΘΦΑΕ‘’’≤ύΊΚΆηΒΡ«ζœΗΨΪΙή÷–Οςœ‘±»Bcl-2ΒΑΑΉΒΡ±μ¥οΝΩ…ΌΘ§“Μ≤ΩΖ÷“‘…Δ‘ΎΒΡΩ≈ΝΘΉ¥¥φ‘Ύ”Ύ…ζΨΪœΗΑϊ÷ ÷–Θ§Νμ“Μ≤ΩΖ÷‘ρΫœ÷¬ΟήΒΊΖ÷≤Φ”Ύ«ζœΗΨΪΙήΒΡΜυΒΉ≤ΩΈΜ(ΆΦ3E)ΓΘ θΚσ3dΈόΟςœ‘±δΜ·Θ§ θΚσ6ΓΪ15dΘ§”ΪΙβΩ≈ΝΘΟςœ‘‘ωΕύΘ§”ΪΙβΟςœ‘‘ω«ΩΘ§“‘8d”»ΈΣΟςœ‘(ΆΦ3F)Θ§“θ–‘Ε‘’’Έ¥Φϊ”ΪΙβ(ΆΦ3FΉσœ¬Ϋ«)ΓΘ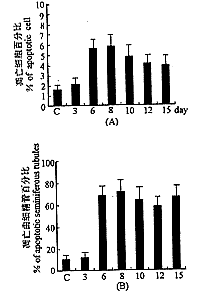
ΆΦ4 ΊΚΆηΒρΆωœΗΑϊ ΐΨίΆ≥ΦΤ(A)ΒρΆωœΗΑϊΑΌΖ÷±»(B)ΒρΆω«ζœΗΨΪΙήΑΌΖ÷±»
Fig.4 Quantitation of testicular apotosis.(A)Percentage of apoptotic cells.(B)Percentage of apoptotic seminiferous tubules.
, http://www.100md.com
Χ÷ ¬έ
≥ΛΤΎ“‘ά¥Θ§»ΥΟ«“Μ÷±‘Ύ―–ΨΩ…ζΨΪœΗΑϊ÷ςΕ·ΆΥΜ·ΒΡΜζ÷ΤΘ§÷ΝΫώ»‘≤Μ °Ζ÷«ε≥ΰΓΘœΗΑϊΒρΆω’β“Μ–¬Η≈ΡνΒΡ≥ωœ÷Θ§ΈΣ―–ΨΩ’ΏΧαΙ©ΝΥ–¬ΥΦ¬ΖΚΆ–¬ΆΨΨΕΓΘ“―”–÷ΛΨί±μΟς…ζΨΪœΗΑϊΒΡΆΥΜ·‘Ύ–ΈΧ§―ß…œΨΏ”–Οςœ‘ΒΡΒρΆωΧΊ’ςΓΘAllanΒ»[1]±®ΒάΨΪ‘≠œΗΑϊΒΡΒρΆω”–3Ηω–ΈΧ§―ßΫΉΕΈΘΚ1.‘γΤΎΘΚœΗΑϊΚΥ»Ψ…Ϊ÷ ΡΐΦ·ΓΔ±ΏΦ·Μ·ΓΘ2.÷–ΤΎΘΚΒρΆω–ΓΧε–Έ≥…≤ΔΩΣ ΦΫΒΫβΓΘ3.ΆμΤΎΘΚΩ…ΦϊΒρΆω–ΓΧεΫΒΫβΒΡΥιΤ§ΓΘΒΪΒρΆωΒΡΤπ ΦΦΑ÷’÷Ι ±ΦδΡ―“‘»ΖΕ®ΓΘ”…”ΎΡΎΖ÷ΟΎΕ‘…ζ÷≥œΗΑϊΒΡΒςΩΊΉς”ΟΘ§–μΕύ Β―ιΧΫΧ÷ΝΥΡΎΖ÷ΟΎΕ‘…ζ÷≥œΗΑϊΒρΆωΒΡΒςΫΎΉς”ΟΓΘTapanainenΒ»[7]±®ΒάΘ§»Ξ¥ΙΧεΈ¥≥… λ¥σ σΊΚΆη…ζΨΪœΗΑϊDNAΫΒΫβΦΑœΗΑϊΒρΆωΟςœ‘‘ωΦ”Θ§GnRHόΉΩΙΦΝ“ύ”–œύΆ§Ής”ΟΘ§ΕχFSHΓΔhCGΦΑΊΚΆΣ‘ρ“÷÷Τ’β÷÷Ής”ΟΘ§“ρ¥Υ»œΈΣ¥Ό–‘œΌΦΛΥΊ «ΒςΫΎΊΚΆη…ζΨΪœΗΑϊ…ζ¥φΚΆΒρΆωΒΡ÷Ί“Σ“ρΥΊΓΘShettyΒ»[8]ΒΡ Β―ι“≤”–άύΥΤΫαΙϊΓΘ
≥ΐ»ΞΡΎΖ÷ΟΎ“ρΥΊΆβΘ§Έ¬Ε»ΓΔΡξΝδΓΔ…ζ≥Λ“ρΉ”ΓΔ…δœΏΓΔΈΔ≤®Β»Εύ÷÷“ρΥΊΨυΕ‘…ζΨΪœΗΑϊΒΡΆΥΜ·”–ΒςΫΎΉς”ΟΓΘ±Ψ Β―ιΦ¥ΩΦ¬«ΒΫΊΚΆη±Ί–κ‘ΎΒΆ”ΎΧεΈ¬4ΓΪ5ΓφΒΡ“θΡ“÷–≤≈Ρή’ΐ≥ΘΖΔ”ΐ’β“ΜΒψΘ§≤…”Ο ÷ θ“ΰΊΚΒΡΖΫΖ®Θ§ΗΡ±δΊΚΆηΖΔ”ΐΜΖΨ≥ΒΡΈ¬Ε»Θ§¥”ΕχΫ®ΝΔ…ζΨΪœΗΑϊΒρΆωΒΡΡΘ–ΆΓΘ’ΐ≥Θ≥…Ρξ–Γ σΒΡΊΚΆη÷–¥φ‘Ύ“ΜΕ® ΐΝΩΒΡΒρΆωœΗΑϊΘ§“‘≥θΦΕΨΪΡΗœΗΑϊΚΆΨΪ‘≠œΗΑϊΈΣ÷ςΘ§ΒΪΒγ”ΨΈόΧί¥χ≥ωœ÷ΓΘ“ΰΊΚ θΚσ6ΓΪ15dΘ§ θ≤ύΊΚΆη÷ΊΝΩœ¬ΫΒΘ§…ζΨΪœΗΑϊΦθ…ΌΘ§ΒρΆωœΗΑϊ‘ωΕύΘ§“‘ΨΪΡΗœΗΑϊΚΆΨΪΉ”œΗΑϊΈΣ÷ςΘ§≤ΔΩ…ΦϊΒρΆω–ΓΧεΘ§Βγ”Ψ”–Οςœ‘ΓΑDNA ladderΓ±≥ωœ÷ΓΘ θΚσ3d‘ρ”κΕ‘’’≤ύΈόΟςœ‘±δΜ·ΓΘ’β“ΜΫαΙϊ”κShikoneΒ»[9]ΒΡ±®Βά”–Υυ≤ΜΆ§ΓΘΥϊΟ«ΒΡ Β―ι÷–Θ§«ύ¥Κ«ΑΤΎ¥σ σΒΡΊΚΆη‘Ύ θΚσ2dΦ¥”–Οςœ‘÷ΊΝΩœ¬ΫΒΘ§…ζΨΪœΗΑϊΦθ…ΌΘ§ΒρΆωœΗΑϊ‘ωΕύΦΑDNAΫΒΫβΓΘΈ“Ο«»œΈΣ’βΩ…Ρή”κ≤ΜΆ§ΡξΝδΕΈΕ‘Έ¬Ε»ΟτΗ––‘≤ΜΆ§ΦΑΕ·Έο÷÷ τ≤ΜΆ§”–ΙΊΓΘΕχHeiskanenΒ»[10]‘Ύ―–ΨΩ»ΥΒΡ“ΰΊΚ ±»¥ΖΔœ÷Θ§ΒρΆω…ζΨΪœΗΑϊ±»’ΐ≥ΘΊΚΆη…ΌΓΘ’β≤ΜΩ…ΡήΆξ»Ϊ «“ρΈΣ÷÷ τΒΡ≤ν±πΓΘ»ΥΒΡ“ΰΊΚ «œ»ΧλΒΡΘ§ΊΚΆη“Μ÷±ΈΜ”ΎΗΙ«Μ37ΓφΒΡΜΖΨ≥÷–Θ§ «¬ΐ–‘ΒΡ≥ΛΤΎΙΐ≥ΧΘ§Εχ Β―ι“ΰΊΚ‘ρ «Α―‘Ύ“θΡ“÷–’ΐ≥ΘΖΔ”ΐΒΡΊΚΆη‘ΌΆΤΜΊΗΙ«ΜΘ§ «Φ±ΥΌΒΡΜΖΨ≥ΗΡ±δΘ§ «“ΜΗωΕΧΤΎΙΐ≥ΧΘ§Υυ“‘Φ¥±ψΕΦ «¥Π”Ύ≤Μ’ΐ≥ΘΒΡΈ¬Ε»ΜΖΨ≥÷–Θ§«ιΩωΒΡ≤ΜΆ§ΜαΒΦ÷¬ΫαΙϊΒΡΆξ»Ϊ≤ΜΆ§ΓΘΈ“Ο«»œΈΣΘ§ Β―ι“ΰΊΚ «Ε‘ΊΚΆηΖΔ”ΐΈ¬Ε»ΜΖΨ≥ΒΡΆΜ»ΜΗΡ±δΘ§Έ¬Ε»ΒΡΒςΫΎΉς”Ο±»œ»Χλ“ΰΊΚΗϋ÷Ί“ΣΗϋΟςœ‘ΓΘΜΙ”–±®ΒάΖΔœ÷“ΰΊΚΜαΒΦ÷¬ΊΚΆΣ≈®Ε»ΒΡœ¬ΫΒ[11]Θ§Ρ«Ο¥Έ¬Ε» «÷±Ϋ”ΒΦ÷¬ΒρΆωΒΡ“ρΥΊΘ§ΜΙ «Ά®ΙΐΦΛΥΊΒςΫΎΒρΆωΘ§ΜρΝΫ’ΏΫ‘”–Θ§”–¥ΐ”ΎΫχ“Μ≤Ϋ―–ΨΩΓΘ
, ΑΌΡ¥“Ϋ“©
bcl-2Μυ“ρ «‘Ύ»ΥBΝήΑΆœΗΑϊΝω÷–±ΜΖΔœ÷ΒΡ“Μ÷÷‘≠Α©Μυ“ρΘ§Bcl-2ΒΑΑΉ «“Μ÷÷ΡΛΒΑΑΉΘ§ΈΜ”ΎœΏΝΘΧεΡΛΓΔΜ§ΟφΡΎ÷ ΆχΡΛΦΑΚΥΡΛ…œ[12]ΓΘBaxΒΑΑΉ «Bcl-2Φ“ΉεΒΡ≥…‘±÷°“ΜΘ§ «Ρή”κBcl-2ΒΑΑΉΫαΚœΒΡΆ§‘¥Ζ÷Ή”Θ§ΝΫ’Ώ‘ΎœΗΑϊ÷–ΒΡ±»άΐΨωΕ®ΝΥœΗΑϊ «ΖώΒρΆωΓΘΒ±Bcl-2ΒΑΑΉ‘ΎœΗΑϊ÷–ΙΐΝΩ±μ¥ο ±Θ§”κΥυ”–BaxΫαΚœΘ§œΗΑϊ≥…ΜνΘ§ΒρΆω“÷÷ΤΘΜΒ±BaxΙΐΝΩ¥φ‘ΎΘ§”κBcl-2–Έ≥…“λ‘¥ΕΰΨέΧεΘ§‘ρœΗΑϊΒρΆω[13]ΓΘΥϋΟ«”κœΗΑϊΒρΆωΒΡΙΊœΒ‘Ύ÷ΉΝωΓΔΝήΑΆœΗΑϊΖΫΟφ“―”–¥σΝΩ―–ΨΩΘ§ΒΪ”κ…ζ÷≥œΗΑϊΒρΆωΒΡΙΊœΒ»¥Κή…Ό±®ΒάΓΘSuzukiΒ»[4]“‘«–≥ΐ–‘œΌΒΡ–έ–‘–Γ σ…ζ÷≥ΙήΒά(«ΑΝ–œΌΓΔΗΫΊΚ)ΈΣΒρΆωΡΘ–ΆΘ§ΖΔœ÷ΥφΉ≈ΒρΆωΒΡ≤ζ…ζ≥ωœ÷Bcl-2œ¬ΒςΘ§≤Δ‘Λ ΨΉ≈fasΜυ“ρΒΡ±μ¥οΓΘΕχΈ“Ο«ΒΡ―–ΨΩ‘ρΖΔœ÷Θ§‘Ύ’ΐ≥Θ–Γ σΊΚΆη…ζΨΪœΗΑϊΒΡΑϊ÷ ÷–”–¥σΝΩBcl-2ΒΑΑΉΓΔ…ΌΝΩBaxΒΑΑΉ±μ¥οΘ§“ΰΊΚ θΚσΒρΆωœΗΑϊΒΡ‘ωΦ”Θ§ΑιΥφΉ≈Bcl-2ΒΑΑΉΒΡΟςœ‘Φθ…ΌΘ§BaxΒΑΑΉΒΡΟςœ‘‘ωΕύΓΘΫαΙϊΧα Ψ‘ΎΊΚΆη…ζΨΪœΗΑϊΒΡΒρΆω÷–Θ§Bcl-2ΚΆBaxΜυ“ρ”–÷Ί“ΣΒςΫΎΉς”ΟΓΘ
ΊΚΆη…ζ÷≥œΗΑϊΒρΆωΦΑœύΙΊΜυ“ρΒςΫΎΉς”ΟΒΡ―–ΨΩΘ§ΈΣΡ––‘…ζ÷≥ΜζάμΒΡΧΫΧ÷ΧαΙ©ΝΥ“ΜΧθ–¬ΆΨΨΕΓΔΕ‘ΫΎ”ΐΓΔ≤Μ”ΐΒ»ΒΡ―–ΨΩΨυ”–÷Ί“Σ“β“εΓΘ
, ΑΌΡ¥“Ϋ“©
ΆΦ3 AΘ§B TUNEL»Ψ…ΪΓΝ300 A.’ΐ≥ΘΊΚΆη«–Τ§ B.“ΰΊΚ θΚσ8dΒΡΊΚΆη«–Τ§ ΦΐΆΖΥυ ΨΈΣ―τ–‘ΕύΚΥΨόœΗΑϊΓΘ(CΓΔDΓΔEΓΔF)…ζΈοΥΊ-ΩΙ…ζΈοΥΊΒΑΑΉDCSΦδΫ”Οβ“Ώ”ΪΙβ»Ψ…Ϊ ΓΝ300
C.Bcl-2ΒΑΑΉ‘Ύ’ΐ≥ΘΊΚΆη÷–ΒΡΖ÷≤Φ D.Bcl-2ΒΑΑΉ‘Ύ“ΰΊΚ θΚσ8dΒΡΊΚΆη÷–ΒΡΖ÷≤Φ E.BaxΒΑΑΉ‘Ύ’ΐ≥ΘΊΚΆη÷–ΒΡΖ÷≤Φ F.BaxΒΑΑΉ‘Ύ“ΰΊΚ θΚσ8dΒΡΊΚΆη÷–ΒΡΖ÷≤Φ Ήσœ¬Ϋ«ΈΣ“θ–‘Ε‘’’
Fig.3 A,B TUNEL staining of the formalin-fixed, paraffin-embedded testicular tissue sections. Postive staining appears green fluorescence. ΓΝ300 A.normal testes. B. cryptorchid testes on day 8. Arrow indicates positive Multinucleated giant cell.
, http://www.100md.com
(C,D,E,F)Biotin-avidin DCS system indirect immunofluorescence staining. Postive staining appears green fluorescence. ΓΝ300 C: Distribution of Bcl-2 protein in normal testes.D: Distribution of Bcl-2 protein in cryptorchid testes on day 8. E: Distribution of Bax protein in normal testes. F: Distribution of Bax protein in cryptorchid testes on day 8. Insert indicates negative control.
* ΙζΦ“Ή‘»ΜΩΤ―ßΜυΫπΉ ÷ζΩΈΧβ(No.39270380)ΚΆΈά…ζ≤ΩΩΤ―–ΜυΫπΉ ÷ζΩΈΧβ(No.94-2-004)
, ΑΌΡ¥“Ϋ“©
Γς Department of Histology and Embryology, Institute of Basic Medical Sciences, CAMS and PUMC, Beijing 10005, China
≤ΈΩΦΈΡœΉ
[1]Allan DJ, Harmon BV, Roberts SA. Spermatogonial apoptosis has three morphologically recognizable phases and shows no circadian rhythm during normal spermatogenesis in the rat. Cell Prolif, 1992,25(3):241
[2]Rodriguez JB, Garcia CM. Spontaneous germ cell death in the testis of the adult rat takes the form of apoptosis: re-evaluation of cell types that exhibit the ability to die during spermatogenesis. Cell Prolif, 1996,29(1):13
, ΑΌΡ¥“Ϋ“©
[3]Ν÷»ξœ…Ήέ ωΘ§Υο÷ΨœΆ…σ–Θ.bcl-2Μυ“ρΦ“Ήε”κœΗΑϊ≥Χ–ρ–‘ΥάΆω.ΙζΆβ“Ϋ―ß“≈¥Ϊ―ßΖ÷≤αΘ§1997Θ§20(5)ΘΚ242
[4]Suzuki A, Matsuzawa A, Iguchi T. Down regulation of bcl-2 is the first step on fas mediated apoptosis of male reproductive tract. Oncogene 1996,13(1):31
[5]Yin YZ, Hawkins KL, Dewolf WC, et al. Heat stress causes testicular germ cell apoptosis in adult mice. J Andro, 1997,18(2),159
[6]Xu ZL, Parker SB, Minkoff R. Distribution of type IV collagen, laminin, and fibronection during maxillary process formation in the chick embryo. Am J Anat, 1990,187(3):232
, ΑΌΡ¥“Ϋ“©
[7]Tapanainen JS, Tilly JL, Vihko KK, et al. Hormonal control of apoptotic cell death in the testis: Gonadotropins and androgens as testicular cell survival factors. Mol Endocrinol, 1993,7(5):643
[8]Shetty J,Marathe GK, Dighe RR, et al. Specific immunoneutralization of FSH leads to apoptotic cell death of the pachytene spermatocytes and spermatogonial cells in the rat. Endocrinol, 1996,137(5):2179
[9]Shikone T, Billig H, Hsueh AJW, et al. Experimentally induced cryptorchidism increases apoptosis in rat testis. Biolo Reproduc, 1994,51(5):865
, http://www.100md.com
[10]Heiskanen P, Billig H, Toppari J, et al. Apoptotic cell death in the normal and cryptorchid human testis: The effect of human chorionic gonadotropin on testicular cell survival. Pediatric Res, 1996,40(2):351
[11]Sharpe RM, Kerr JB, Fraser HM, et al. Intratesticular factors and testosterone secretion: effect of treatments that alter the level of testosterone within the testis. J Androl, 1986, 7(3):180
[12]Nguyen M, Branton PE, Walton PA, et al. Role of membrane anchor domain of bcl-2 in suppression of apoptosis caused by E1B-defective adenovirus. J Biol Chem, 1994,269(24):16521
[13]Oltrai ZN, Milliman CL, Korsmeyer SJ, et al. Bcl-2 heterodimerizes in vivo with a conserved homolog, Bax, that accelerates programmed cell death. Cell, 1993,74(4):609
’Ηε1998-01 –όΜΊ1998-05, ΑΌΡ¥“Ϋ“©
ΒΞΈΜΘΚ–λΫΓ –μ‘ω¬Μ ΫΣ”ΔΧΈ «°œΰίΦ ΜΤ”ώήΏ(÷–Ιζ“Ϋ―ßΩΤ―ß‘Κ ÷–Ιζ–≠ΚΆ“ΫΩΤ¥σ―ßΜυ¥Γ“Ϋ―ß―–ΨΩΥυΘ§±±Ψ© 100005)
ΙΊΦϋ¥ ΘΚ…ζΨΪœΗΑϊΒρΆωΘΜBcl-2ΒΑΑΉΘΜBaxΒΑΑΉ
ΫβΤ ―ß±®990121
ΓΨ’Σ“ΣΓΩ ΡΩΒΡ ―–ΨΩ ÷ θ“ΰΊΚΥυ÷¬≥…Ρξ–Γ σ…ζ÷≥œΗΑϊΒρΆωΦΑœύΙΊΒςΫΎ“ρΉ”bcl-2ΓΔbaxΒΑΑΉ‘Ύ«ζœΗΨΪΙή÷–ΒΡΕ®ΈΜΓΔ±δΜ·ΓΘ ΖΫΖ® “‘Ρ©ΕΥΆ――θΚΥΧ«ΚΥΥαΉΣ“ΤΟΗΫιΒΦΒΡdUTP»±ΩΎΡ©ΕΥ±ξΦ«Ζ®(terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling method,TUNEL)Φλ≤βΒρΆωΒΡ…ζ÷≥œΗΑϊΘ§…ζΈοΥΊ-ΩΙ…ζΈοΥΊΒΑΑΉDCS(biotin-avidin D cell sorter)ΧεœΒΦδΫ”Οβ“Ώ”ΪΙβΖ®Φλ≤βBcl-2ΓΔBaxΒΑΑΉ‘Ύ«ζœΗΨΪΙή÷–ΒΡΕ®ΈΜΓΔΖ÷≤ΦΓΘ ΫαΙϊ “ΰΊΚ θΚσ3dΘ§ ÷ θ≤ύΊΚΆη÷ΊΝΩΦΑœΗΑϊΒρΆω ΐΝΩ”κΕ‘≤ύΈόΟςœ‘«χ±πΓΘΕχ6ΓΪ15dΘ§ΊΚΆη÷ΊΝΩΟςœ‘Φθ«αΘ§ΒρΆωœΗΑϊΟςœ‘‘ωΕύΘ§“‘ΨΪΡΗœΗΑϊΓΔΨΪΉ”œΗΑϊΈΣ÷ςΓΘ‘Ύ’ΐ≥Θ–Γ σ«ζœΗΨΪΙή÷–Θ§bcl-2¥σΝΩ±μ¥οΘ§≥ …Δ‘ΎΒΡΩ≈ΝΘΉ¥¥φ‘Ύ”ΎΗςΦΕ…ζΨΪœΗΑϊ÷–Θ§“‘ΨΪΉ”œΗΑϊΑϊ÷ ΈΣ÷ςΓΘBaxΒΑΑΉ±μ¥οΝΩΫœ…ΌΘ§“Μ≤ΩΖ÷“≤“‘…Δ‘ΎΒΡΩ≈ΝΘ¥φ‘Ύ”ΎΗςΦΕ…ζΨΪœΗΑϊ÷ ÷–Θ§Νμ“Μ≤ΩΖ÷‘ρΫœ÷¬ΟήΒΊΖ÷≤Φ”Ύ«ζœΗΨΪΙήΜυΒΉ≤ΩΓΘ“ΰΊΚ θΚσ3dΘ§Bcl-2ΓΔBaxΒΡ±μ¥ο”κΕ‘≤ύΈόΟςœ‘«χ±πΘΜ6ΓΪ15dΘ§Bcl-2Οςœ‘œ¬ΫΒΘ§BaxΟςœ‘‘ωΦ”ΓΘ Ϋα¬έ “ΰΊΚΟςœ‘”’ΒΦ…ζ÷≥œΗΑϊΒρΆωΘ§Bcl-2ΓΔBaxΒΑΑΉΕ‘…ζ÷≥œΗΑϊΒΡΒρΆω”–÷Ί“ΣΒςΫΎΉς”ΟΘ§Χα ΨΈ¬Ε»ΓΔBcl-2ΓΔBaxΒΑΑΉ‘ΎΨΪΉ”ΖΔ…ζΙΐ≥Χ÷–Ε‘…ζΨΪœΗΑϊΒΡ÷ςΕ·ΆΥΜ·Ω…Ρή”–÷Ί“ΣΉς”ΟΓΘ
, ΑΌΡ¥“Ϋ“©
CRYPTORCHIDISM INDUCES TESTICULAR GERM CELL APOPTOSIS
IN MICE TESTES AND CHANGES OF Bcl-2,Bax PROTEIN EXPRESSION
Xu Jian, Xu ZengluΓς, Jiang Yingtao, Qian Xiaojing, Huang Yuling
(Institute of Basic Medical Sciences, CAMS and PUMC, Beijing)
ΓΨAbstractΓΩ Objective To investigate the effect of experimentally induced cryptorchidism on apoptosis in testes of adult mice and the distribution changes of Bcl-2, Bax protein. Methods Total DNA was isolated from testes and was electrophoretically separated in 1.5% agarose gel, TUNEL method, biotin-avidin DCS system indirect immunofluorescence techniques were used. Results DNA fragmentation, increasing apoptotic germ cells, in coordination with the loss of testicular weight, were observed on 6-15d in the cryptorchid testes. By TUNEL staining it was noted that postive cells were most frequently among primary spermatocytes and round spermatids. Bcl-2, Bax protein mainly distributed in cytoplasm of germ cells. With the increase of apoptosis, Bcl-2 was down and Bax was up regulated. Conclusion Temperature, Bcl-2, and Bax protein play an important role in male germ cell apoptosis.
, ΑΌΡ¥“Ϋ“©
ΓΨKey wordsΓΩ Apoptotic germ cells; Bcl-2 protein; Bax protein
’ΐ≥ΘΒΡ…ζΨΪΙΐ≥Χ÷–Θ§¥φ‘ΎΉ≈…ζ÷≥œΗΑϊΒΡ÷ςΕ·ΆΥΜ·œ÷œσΘ§≥ΛΤΎ“‘ά¥ ήΒΫ―–ΨΩ’ΏΒΡ÷Ί ”Θ§ΤδΜζ÷Τ»‘≤Μ«ε≥ΰΓΘΫϋΦΗΡξά¥Θ§ΙζΆβ“Μ–©―–ΨΩ’ΏΩΣ ΦΫΪ’β÷÷÷ςΕ·ΆΥΜ·”κœΗΑϊΒρΆωΝΣœΒ‘Ύ“ΜΤπΘ§ΚήΕύ Β―ι“≤±μΟςΒρΆωΩ…Ρή «…ζ÷≥œΗΑϊ÷ςΕ·ΆΥΜ·ΒΡ“Μ÷÷Μζ÷Τ[1Θ§2]ΓΘ”…”Ύœ¬«πΡ‘-¥ΙΧε-–‘œΌ÷αΕ‘…ζ÷≥ΒΡΟςœ‘ΩΊ÷ΤΉς”ΟΘ§¥σΕύ ΐ Β―ιΦ·÷–”Ύ”–ΙΊΡΎΖ÷ΟΎΕ‘…ζ÷≥œΗΑϊΒρΆωΒςΫΎΖΫΟφΒΡ―–ΨΩΘ§ΕχΊΚΆηΜΙ±Ί–κ‘ΎΒΆ”ΎΧεΈ¬4ΓΪ5ΓφΒΡ“θΡ“÷–≤≈Ρή’ΐ≥ΘΖΔ”ΐΘ§Έ¬Ε»“≤ «÷Ί“ΣΒΡΒςΫΎ“ρΥΊ÷°“ΜΓΘ“ρ¥ΥΘ§Έ“Ο«“‘ΒΞ≤ύ ÷ θ“ΰΊΚΗΡ±δΨΪΉ”ΖΔ…ζΥυ–η“ΣΒΡ’ΐ≥ΘΈ¬Ε»ΒΡΖΫΖ®ά¥Ϋ®ΝΔ…ζ÷≥œΗΑϊΒρΆωΒΡΡΘ–ΆΓΘ
–μΕύ―–ΨΩ±μΟςbcl-2Μυ“ρΦ“Ήε «ΒςΫΎœΗΑϊΒρΆωΒΡ“ΜΗω÷Ί“ΣΜυ“ρΦ“ΉεΘ§ΒΪΦ“Ήε≥…‘±Ε‘ΒρΆωΒΡΉς”Ο»¥”–Οςœ‘≤ΜΆ§ΓΘ“Μάύ¥ΌΫχœΗΑϊΒρΆω»γbaxΓΔbcl-xsΒ»Θ§Νμ“Μάύ‘ρ“÷÷ΤΒρΆω»γbcl-2ΓΔbcl-xΒ»[3]ΓΘbcl-2Μυ“ρΦ“ΉεΕ‘÷ΉΝωœΗΑϊΓΔΝήΑΆœΗΑϊΒρΆωΉς”ΟΒΡ―–ΨΩΚήΕύΘ§Ε‘…ζ÷≥œΗΑϊΒρΆωΒΡΉς”ΟΘ§ΉνΫϋ“≤”–Ηω±π±®Βά[4]ΓΘΈ“Ο«‘ρ―–ΨΩΝΥBcl-2ΓΔBaxΒΑΑΉ‘Ύ’ΐ≥Θ–Γ σΊΚΆη«ζœΗΨΪΙή÷–ΒΡΕ®ΈΜΦΑΖ÷≤ΦΘ§ΦΑΤδ‘Ύ…ζΨΪœΗΑϊΒρΆω ±Θ§±μ¥οΒΡΗΡ±δΓΘ
, http://www.100md.com
≤ΡΝœΚΆΖΫΖ®
1.Ε·Έοά¥‘¥ΦΑΕ·ΈοΡΘ–ΆΒΡΫ®ΝΔ
÷–Ιζ“Ϋ―ßΩΤ―ß‘ΚΕ·ΈοΥυΧαΙ©ΒΡ8ΓΪ10÷ή(26ΓΪ30g)ΫΓΩΒ–έ–‘άΞΟς–Γ σΓΘΈλΑΆ±»ΆΉΗΙ«ΜΉΔ…δ¬ιΉμ(30mg/kg)Θ§ΖλΚœΒΞ≤ύΗΙΙ…ΙΒΙήΘ§ Ι–Γ σΊΚΆη≤ΜΡή¥”ΗΙ«Μœ¬ΫΒ÷Ν“θΡ“Θ§Έ¥ ÷ θ≤ύΦΑ’ΐ≥Θ–Γ σΒΡΊΚΆηΉςΕ‘’’Θ§ θΚσ3ΓΔ6ΓΔ8ΓΔ10ΓΔ12ΓΔ15d»Γ≤ΡΘ§ΟΩΉιΕ·Έο6ΓΪ8÷ΜΓΘ
2. ‘ΦΝ
œΗΑϊΒρΆω ‘ΦΝΚ–(ClontechΙΪΥΨ)Θ§ΆΟΩΙ–Γ σBcl-2ΓΔBaxΕύΩΙ(Santa Cruz BiotechnologyΙΪΥΨΘ§ΙΚ”Ύ±±Ψ©÷–…Ϋ…ζΈοΦΦ θ”–œόΙΪΥΨ)Θ§…ζΈοΥΊΜ·ΒΡ…Ϋ―ρΩΙΆΟIgGΘ§”ΪΙβΥΊ-ΩΙ…ζΈοΥΊΒΑΑΉDCSΓΔ…Ϋ―ρ―Σ«ε(Vector Laboratories, Burlingame),ppd(phenylenediamine)ΓΔagaroseΓΔRNase AΓΔprotein kinase (SigmaΙΪΥΨ)ΓΘ
, http://www.100md.com
3.ΊΚΆη≥Τ÷ΊΦΑHE»Ψ…Ϊ
–Γ σΨ±ΉΒΆ―Ψ Ζ®¥ΠΥάΘ§»ΓΥΪ≤ύΊΚΆηΘ§≥Τ÷ΊΓΘ10%÷––‘ΗΘΕϊ¬μΝ÷ΙΧΕ®24hΘ§≥ΘΙφΑϋ¬ώΓΔ«–Τ§ΓΔHE»Ψ…ΪΓΘ
4.Ήι÷·DNAΧα»ΓΦΑ«μ÷§Χ«ΡΐΫΚΒγ”Ψ
¥”¥ΠΥάΒΡ–Γ σ÷–»Γ≥ωΊΚΆη(Ω…-70Γφ±Θ¥φ)Θ§»ΞΑΉΡΛΘ§―–ΥιΘ§Φ”»κ10±ΕΧεΜΐΒΡbuffer(10mmol/L Tris-HCl, pH8.0; 0.1mol/L EDTA,pH8.0;0.5%SDS; 100mmol/L NaCl,4ΓφΙΐ“Ι)Θ§Φ”RNase A÷Ν÷’≈®Ε»40mg/L,37ΓφΘ§1hΘΜΦ”protein kinase÷Ν÷’≈®Ε»100mg/L,50ΓφΘ§3h;10 000gάκ–Ρ30minΘ§…œ«ε”ΟΖ”/¬»Ζ¬≥ιΧα2¥ΈΘ§ΈόΥ°““¥Φ-20ΓφΙΐ“ΙΘ§12 000gάκ–Ρ30minΓΘ’φΩ’≥ιΗ…Θ§TE buffer (10mmol/L Tris HCl pH8.0; 1mmol/L EDTA, pH8.0)»ήΫβΓΘ1.5%«μ÷§Χ«ΡΐΫΚΒγ”ΨΓΘ
, ΑΌΡ¥“Ϋ“©
5.ΒρΆωœΗΑϊΒΡΦλ≤βΦΑΕ®ΝΩ ΐΨί¥Πάμ
Α¥’’ ‘ΦΝΚ–ΥΒΟςΘ§“‘TUNELΖ®Φλ≤βΒρΆωœΗΑϊΘ§œ‘¬Χ…Ϊ”ΪΙβΒΡœΗΑϊΈΣ―τ–‘œΗΑϊΓΘΑ¥Yin[5]Ζ®Ής ΐΨί¥ΠάμΘΚœ‘ΈΔΨΒœ¬»Έ―Γ5ΓΪ10Ηω ”“ΑΘ§ΦΤ ΐ50Ηω«ζœΗΨΪΙήΚα«–ΟφΘ§Κ§ΒρΆωœΗΑϊ’ΏΈΣ―τ–‘«ζœΗΨΪΙήΘ§‘ΌΦΤ ΐ15ΓΪ20Ηω―τ–‘«ζœΗΨΪΙή÷–ΒΡΉήœΗΑϊ ΐΚΆ―τ–‘œΗΑϊ ΐΘ§ΦΤΥψΒρΆωœΗΑϊΒΡΑΌΖ÷±»ΓΘΟΩΗω ±ΦδΒψΦΤΥψ3÷Μ–Γ σΒΡΊΚΆηΘ§ΟΩΗωΊΚΆη ΐ3’≈«–Τ§ΓΘ
6.…ζΈοΥΊ-ΩΙ…ζΈοΥΊΒΑΑΉDCSΦδΫ”Οβ“Ώ”ΪΙβ»Ψ…Ϊ
»Γ–¬œ ΊΚΆηΉι÷·Θ§OCTΑϋ¬ώΘ§“ΚΒΣ÷–άδΕ≥Θ§Κψάδœδ«–Τ§Θ§Κώ10ΠΧmΘ§0.1mol/L PBS≈δ÷ΤΒΡ10%…Ϋ―ρ―Σ«ε4ΓφΖθ”ΐ20minΘ§¬Υ÷ΫΈϋ»ΞΕύ”ύ―Σ«εΘ§Φ”…œ”Ο10%…Ϋ―ρ―Σ«εœΓ ΆΒΡ(1ΓΟ100)ΆΟΩΙ–Γ σBcl-2ΓΔBaxΕύΩΙΘ§4ΓφΙΐ“ΙΘ§“θ–‘Ε‘’’ ûΥΥ≤Ϋ÷ηΘ§άδPBS 4Γφœ¥3¥ΈΘ§ΟΩ¥Έ5minΘ§Φ”…œ10%…Ϋ―ρ―Σ«εœΓ ΆΒΡ(1ΓΟ200)…ζΈοΥΊΜ·…Ϋ―ρΩΙΆΟIgGΘ§4ΓφΖθ”ΐ1hΘ§άδPBS”Ύ4Γφœ¥3¥ΈΘ§ΟΩ¥Έ5minΓΘ“‘”ΪΙβΥΊ-ΩΙ…ζΈοΥΊΒΑΑΉDCS (1ΓΟ200ΧΦΥα«βΡΤΜΚ≥ε“ΚœΓ Ά)4ΓφΖθ”ΐ1hΘ§άδPBS 4Γφœ¥4¥ΈΘ§ΟΩ¥Έ5minΘ§”ΟΚ§10%PBSΚΆ1% ppdΒΡΗ ”ΆΖβΤ§[6]ΓΘ±ξ±Ψ‘ΎOlympusΖ¥…δ Ϋ”ΪΙβœ‘ΈΔΨΒœ¬Ιέ≤λΘ§≤Δ”Ο400 ASA KodakΫΚΨμ…ψΤ§ΓΘ
, http://www.100md.com
Ϋα Ιϊ
1.ΊΚΆη÷ΊΝΩΒΡΗΡ±δ
’ΐ≥Θ σΦΑΕ‘’’≤ύΊΚΆηΒΡ÷ΊΝΩ‘Ύ’ϊΗω Β―ιΙΐ≥Χ÷–ΈόΟςœ‘±δΜ·ΓΘ“ΰΊΚ÷ΊΝΩ‘Ύ θΚσ3d”κΕ‘’’≤ύΈόΟςœ‘±δΜ·ΓΘ θΚσ6dΦθ«α20%Θ§8dΦθ«α29%Θ§10dΦθ«α32%Θ§12dΦθ«α46%Θ§15dΦθ«α31%ΓΘ“ΰΊΚ≤ύ”κΕ‘’’≤ύ÷ΊΝΩΒΡ≤ν±π‘Ύ6ΓΪ15dΟΩΗω ±ΦδΒψΨυ”–œ‘÷χ≤ν“λ(P<0.01)Θ§ΕχΉιΦδΈόœ‘÷χ≤ν“λ(ΆΦ1)ΓΘ
ΆΦ1 “ΰΊΚΦΑΕ‘’’≤ύΊΚΆη÷ΊΝΩΒΡ±δΜ·
Fig.1 Both cryptorchid and control testesweights (
) are plotted against days of experimental cryptorchidism from 3 to 15., http://www.100md.com
2.HE»Ψ…Ϊ
Ε‘’’≤ύΊΚΆη…ζΨΪ…œΤΛΆξ’ϊΘ§ΗςΦΕ…ζΨΪœΗΑϊΦΑΨΪΉ”≈≈Ν–”––ρΓΘ“ΰΊΚ θΚσ3dΈόΟςœ‘±δΜ·ΓΘ θΚσ6ΓΪ15d«ζœΗΨΪΙήΙή«Μ‘ω¥σΘ§…ζΨΪ…œΤΛ±δ±ΓΘ§≤ΜΆξ’ϊΘ§ΨΪΉ”Οςœ‘Φθ…ΌΘ§≤ΔΩ…Φϊ“Μ–©œΗΑϊΒΡ»Ψ…Ϊ÷ Οςœ‘±ΏΦ·Μ·Θ§≥ ΜΖΉ¥ΓΘ“Μ–©«ζœΗΨΪΙή÷–≥ωœ÷ΓΑΕύΚΥΨόœΗΑϊΓ±(multinucleated giant cells)Θ§Φ¥ΕύΗωœΗΑϊΚΥΨέΦ·–Έ≥…“ΜΗω¥σΒΡœΗΑϊΆ≈ΩιΘ§Τδ÷–“Μ–©œΗΑϊΚΥ»Ψ…Ϊ÷ Οςœ‘±ΏΦ·Μ·Θ§≥ ΜΖΉ¥ΓΘΦδ÷ œΗΑϊΚΆ÷ß≥÷œΗΑϊΈό±δΜ·ΓΘ
3.ΡΐΫΚΒγ”Ψ
“ΰΊΚ θΚσ6ΓΔ8ΓΔ10dΩ…ΦϊΓΑDNA ladderΓ±ΆΦœώΘ§œ‘ Ψ”–ΚΥ–ΓΧεΦδDNAΕœΝ―ΒΡ–ΓΤ§ΕΈ¥φ‘ΎΘ§Εχœύ”ΠΒΡΕ‘’’≤ύΓΔ“ΰΊΚ θΚσ3dΦΑ’ΐ≥Θ–Γ σΊΚΆηΟΜ”–Ιέ≤λΒΫ–ΓΤ§ΕΈDNA(ΆΦ2)ΓΘ
ΆΦ2 DNA«μ÷§Χ«ΡΐΫΚΒγ”ΨΆΦΤΉ
, http://www.100md.com
Fig.2 Agarose gel fractionation of DNA.Low molecular-weight
DNA from both cryptorchid and control testes were fractionated
on 1.5% agarose gel containing 0.5mg/L ethidium bromide.
4.TUNEL»Ψ…Ϊ
’ΐ≥ΘΦΑΕ‘’’≤ύΊΚΆη”–…Ό ΐ«ζœΗΨΪΙή÷–Ω…Ιέ≤λΒΫΗω±πΒρΆωœΗΑϊΘ§÷ς“ΣΈΣ≥θΦΕΨΪΡΗœΗΑϊ(ΆΦ3A)ΓΘ“ΰΊΚ θΚσ3dΈόΟςœ‘±δΜ·Θ§ θΚσ6ΓΪ15d―τ–‘«ζœΗΨΪΙήΦΑ―τ–‘œΗΑϊΨυΟςœ‘‘ωΦ”ΓΘΒρΆωœΗΑϊ÷ς“ΣΑϋά®ΨΪΡΗœΗΑϊΚΆΨΪΉ”œΗΑϊΘ§≤ΩΖ÷ΓΑΕύΚΥΨόœΗΑϊΓ±“≤≥ ―τ–‘ΓΘΈ¥ΦϊΦδ÷ œΗΑϊΦΑ÷ß≥÷œΗΑϊΒρΆω(ΆΦ3B)ΓΘ’ΐ≥ΘΦΑΕ‘’’≤ύΊΚΆη―τ–‘«ζœΗΨΪΙή(Κ§ΒρΆωœΗΑϊΒΡ«ζœΗΨΪΙή)ΈΣ10.7%Θ§―τ–‘œΗΑϊ(ΒρΆωœΗΑϊ)’Φ1.6%ΓΘ“ΰΊΚ θΚσ3d―τ–‘«ζœΗΨΪΙή12%Θ§―τ–‘œΗΑϊ2.1%ΓΘ θΚσ6d―τ–‘«ζœΗΨΪΙή68.6%Θ§―τ–‘œΗΑϊ5.6%Θ§ θΚσ8d―τ–‘«ζœΗΨΪΙή72%Θ§―τ–‘œΗΑϊ5.8%Θ§ θΚσ10d―τ–‘«ζœΗΨΪΙή64.6%Θ§―τ–‘œΗΑϊ4.8%Θ§ θΚσ12d―τ–‘«ζœΗΨΪΙή58.9%Θ§―τ–‘œΗΑϊ4.1%Θ§ θΚσ15d―τ–‘«ζœΗΨΪΙή67.2%Θ§―τ–‘œΗΑϊ3.9%(ΆΦ4AΘ§B)ΓΘ θΚσ3d”κΕ‘’’≤ύΈόΟςœ‘≤ν“λ(P>0.05)ΓΘ θΚσ6ΓΪ15d”κΕ‘’’≤ύΦΑ3dΨυ”–œ‘÷χ≤ν“λ(P<0.05)Θ§6ΓΪ15dΗςΉι÷°ΦδΈόœ‘÷χ≤ν“λ(P>0.05)ΓΘ
, ΑΌΡ¥“Ϋ“©
5.Bcl-2ΓΔBaxΒΑΑΉ‘Ύ«ζœΗΨΪΙή÷–ΒΡΖ÷≤ΦΦΑ±δΜ·
‘Ύ’ΐ≥ΘΦΑΕ‘’’≤ύΊΚΆηΒΡ«ζœΗΨΪΙή÷–Θ§Bcl-2ΒΑΑΉ“‘…Δ‘ΎΒΡΩ≈ΝΘΉ¥¥σΝΩΖ÷≤Φ”ΎΗςΦΕ…ζΨΪœΗΑϊ÷–Θ§“‘ΨΪΉ”œΗΑϊ÷ ΈΣΉνΕύ(ΆΦ3C)Θ§ θΚσ3dΈόΟςœ‘±δΜ·Θ§ θΚσ6ΓΪ15d‘ρΦϊ”ΪΙβΩ≈ΝΘΟςœ‘Φθ…Ό(ΆΦ3D)ΓΘΕχBaxΒΑΑΉ‘Ύ’ΐ≥ΘΦΑΕ‘’’≤ύΊΚΆηΒΡ«ζœΗΨΪΙή÷–Οςœ‘±»Bcl-2ΒΑΑΉΒΡ±μ¥οΝΩ…ΌΘ§“Μ≤ΩΖ÷“‘…Δ‘ΎΒΡΩ≈ΝΘΉ¥¥φ‘Ύ”Ύ…ζΨΪœΗΑϊ÷ ÷–Θ§Νμ“Μ≤ΩΖ÷‘ρΫœ÷¬ΟήΒΊΖ÷≤Φ”Ύ«ζœΗΨΪΙήΒΡΜυΒΉ≤ΩΈΜ(ΆΦ3E)ΓΘ θΚσ3dΈόΟςœ‘±δΜ·Θ§ θΚσ6ΓΪ15dΘ§”ΪΙβΩ≈ΝΘΟςœ‘‘ωΕύΘ§”ΪΙβΟςœ‘‘ω«ΩΘ§“‘8d”»ΈΣΟςœ‘(ΆΦ3F)Θ§“θ–‘Ε‘’’Έ¥Φϊ”ΪΙβ(ΆΦ3FΉσœ¬Ϋ«)ΓΘ
ΆΦ4 ΊΚΆηΒρΆωœΗΑϊ ΐΨίΆ≥ΦΤ(A)ΒρΆωœΗΑϊΑΌΖ÷±»(B)ΒρΆω«ζœΗΨΪΙήΑΌΖ÷±»
Fig.4 Quantitation of testicular apotosis.(A)Percentage of apoptotic cells.(B)Percentage of apoptotic seminiferous tubules.
, http://www.100md.com
Χ÷ ¬έ
≥ΛΤΎ“‘ά¥Θ§»ΥΟ«“Μ÷±‘Ύ―–ΨΩ…ζΨΪœΗΑϊ÷ςΕ·ΆΥΜ·ΒΡΜζ÷ΤΘ§÷ΝΫώ»‘≤Μ °Ζ÷«ε≥ΰΓΘœΗΑϊΒρΆω’β“Μ–¬Η≈ΡνΒΡ≥ωœ÷Θ§ΈΣ―–ΨΩ’ΏΧαΙ©ΝΥ–¬ΥΦ¬ΖΚΆ–¬ΆΨΨΕΓΘ“―”–÷ΛΨί±μΟς…ζΨΪœΗΑϊΒΡΆΥΜ·‘Ύ–ΈΧ§―ß…œΨΏ”–Οςœ‘ΒΡΒρΆωΧΊ’ςΓΘAllanΒ»[1]±®ΒάΨΪ‘≠œΗΑϊΒΡΒρΆω”–3Ηω–ΈΧ§―ßΫΉΕΈΘΚ1.‘γΤΎΘΚœΗΑϊΚΥ»Ψ…Ϊ÷ ΡΐΦ·ΓΔ±ΏΦ·Μ·ΓΘ2.÷–ΤΎΘΚΒρΆω–ΓΧε–Έ≥…≤ΔΩΣ ΦΫΒΫβΓΘ3.ΆμΤΎΘΚΩ…ΦϊΒρΆω–ΓΧεΫΒΫβΒΡΥιΤ§ΓΘΒΪΒρΆωΒΡΤπ ΦΦΑ÷’÷Ι ±ΦδΡ―“‘»ΖΕ®ΓΘ”…”ΎΡΎΖ÷ΟΎΕ‘…ζ÷≥œΗΑϊΒΡΒςΩΊΉς”ΟΘ§–μΕύ Β―ιΧΫΧ÷ΝΥΡΎΖ÷ΟΎΕ‘…ζ÷≥œΗΑϊΒρΆωΒΡΒςΫΎΉς”ΟΓΘTapanainenΒ»[7]±®ΒάΘ§»Ξ¥ΙΧεΈ¥≥… λ¥σ σΊΚΆη…ζΨΪœΗΑϊDNAΫΒΫβΦΑœΗΑϊΒρΆωΟςœ‘‘ωΦ”Θ§GnRHόΉΩΙΦΝ“ύ”–œύΆ§Ής”ΟΘ§ΕχFSHΓΔhCGΦΑΊΚΆΣ‘ρ“÷÷Τ’β÷÷Ής”ΟΘ§“ρ¥Υ»œΈΣ¥Ό–‘œΌΦΛΥΊ «ΒςΫΎΊΚΆη…ζΨΪœΗΑϊ…ζ¥φΚΆΒρΆωΒΡ÷Ί“Σ“ρΥΊΓΘShettyΒ»[8]ΒΡ Β―ι“≤”–άύΥΤΫαΙϊΓΘ
≥ΐ»ΞΡΎΖ÷ΟΎ“ρΥΊΆβΘ§Έ¬Ε»ΓΔΡξΝδΓΔ…ζ≥Λ“ρΉ”ΓΔ…δœΏΓΔΈΔ≤®Β»Εύ÷÷“ρΥΊΨυΕ‘…ζΨΪœΗΑϊΒΡΆΥΜ·”–ΒςΫΎΉς”ΟΓΘ±Ψ Β―ιΦ¥ΩΦ¬«ΒΫΊΚΆη±Ί–κ‘ΎΒΆ”ΎΧεΈ¬4ΓΪ5ΓφΒΡ“θΡ“÷–≤≈Ρή’ΐ≥ΘΖΔ”ΐ’β“ΜΒψΘ§≤…”Ο ÷ θ“ΰΊΚΒΡΖΫΖ®Θ§ΗΡ±δΊΚΆηΖΔ”ΐΜΖΨ≥ΒΡΈ¬Ε»Θ§¥”ΕχΫ®ΝΔ…ζΨΪœΗΑϊΒρΆωΒΡΡΘ–ΆΓΘ’ΐ≥Θ≥…Ρξ–Γ σΒΡΊΚΆη÷–¥φ‘Ύ“ΜΕ® ΐΝΩΒΡΒρΆωœΗΑϊΘ§“‘≥θΦΕΨΪΡΗœΗΑϊΚΆΨΪ‘≠œΗΑϊΈΣ÷ςΘ§ΒΪΒγ”ΨΈόΧί¥χ≥ωœ÷ΓΘ“ΰΊΚ θΚσ6ΓΪ15dΘ§ θ≤ύΊΚΆη÷ΊΝΩœ¬ΫΒΘ§…ζΨΪœΗΑϊΦθ…ΌΘ§ΒρΆωœΗΑϊ‘ωΕύΘ§“‘ΨΪΡΗœΗΑϊΚΆΨΪΉ”œΗΑϊΈΣ÷ςΘ§≤ΔΩ…ΦϊΒρΆω–ΓΧεΘ§Βγ”Ψ”–Οςœ‘ΓΑDNA ladderΓ±≥ωœ÷ΓΘ θΚσ3d‘ρ”κΕ‘’’≤ύΈόΟςœ‘±δΜ·ΓΘ’β“ΜΫαΙϊ”κShikoneΒ»[9]ΒΡ±®Βά”–Υυ≤ΜΆ§ΓΘΥϊΟ«ΒΡ Β―ι÷–Θ§«ύ¥Κ«ΑΤΎ¥σ σΒΡΊΚΆη‘Ύ θΚσ2dΦ¥”–Οςœ‘÷ΊΝΩœ¬ΫΒΘ§…ζΨΪœΗΑϊΦθ…ΌΘ§ΒρΆωœΗΑϊ‘ωΕύΦΑDNAΫΒΫβΓΘΈ“Ο«»œΈΣ’βΩ…Ρή”κ≤ΜΆ§ΡξΝδΕΈΕ‘Έ¬Ε»ΟτΗ––‘≤ΜΆ§ΦΑΕ·Έο÷÷ τ≤ΜΆ§”–ΙΊΓΘΕχHeiskanenΒ»[10]‘Ύ―–ΨΩ»ΥΒΡ“ΰΊΚ ±»¥ΖΔœ÷Θ§ΒρΆω…ζΨΪœΗΑϊ±»’ΐ≥ΘΊΚΆη…ΌΓΘ’β≤ΜΩ…ΡήΆξ»Ϊ «“ρΈΣ÷÷ τΒΡ≤ν±πΓΘ»ΥΒΡ“ΰΊΚ «œ»ΧλΒΡΘ§ΊΚΆη“Μ÷±ΈΜ”ΎΗΙ«Μ37ΓφΒΡΜΖΨ≥÷–Θ§ «¬ΐ–‘ΒΡ≥ΛΤΎΙΐ≥ΧΘ§Εχ Β―ι“ΰΊΚ‘ρ «Α―‘Ύ“θΡ“÷–’ΐ≥ΘΖΔ”ΐΒΡΊΚΆη‘ΌΆΤΜΊΗΙ«ΜΘ§ «Φ±ΥΌΒΡΜΖΨ≥ΗΡ±δΘ§ «“ΜΗωΕΧΤΎΙΐ≥ΧΘ§Υυ“‘Φ¥±ψΕΦ «¥Π”Ύ≤Μ’ΐ≥ΘΒΡΈ¬Ε»ΜΖΨ≥÷–Θ§«ιΩωΒΡ≤ΜΆ§ΜαΒΦ÷¬ΫαΙϊΒΡΆξ»Ϊ≤ΜΆ§ΓΘΈ“Ο«»œΈΣΘ§ Β―ι“ΰΊΚ «Ε‘ΊΚΆηΖΔ”ΐΈ¬Ε»ΜΖΨ≥ΒΡΆΜ»ΜΗΡ±δΘ§Έ¬Ε»ΒΡΒςΫΎΉς”Ο±»œ»Χλ“ΰΊΚΗϋ÷Ί“ΣΗϋΟςœ‘ΓΘΜΙ”–±®ΒάΖΔœ÷“ΰΊΚΜαΒΦ÷¬ΊΚΆΣ≈®Ε»ΒΡœ¬ΫΒ[11]Θ§Ρ«Ο¥Έ¬Ε» «÷±Ϋ”ΒΦ÷¬ΒρΆωΒΡ“ρΥΊΘ§ΜΙ «Ά®ΙΐΦΛΥΊΒςΫΎΒρΆωΘ§ΜρΝΫ’ΏΫ‘”–Θ§”–¥ΐ”ΎΫχ“Μ≤Ϋ―–ΨΩΓΘ
, ΑΌΡ¥“Ϋ“©
bcl-2Μυ“ρ «‘Ύ»ΥBΝήΑΆœΗΑϊΝω÷–±ΜΖΔœ÷ΒΡ“Μ÷÷‘≠Α©Μυ“ρΘ§Bcl-2ΒΑΑΉ «“Μ÷÷ΡΛΒΑΑΉΘ§ΈΜ”ΎœΏΝΘΧεΡΛΓΔΜ§ΟφΡΎ÷ ΆχΡΛΦΑΚΥΡΛ…œ[12]ΓΘBaxΒΑΑΉ «Bcl-2Φ“ΉεΒΡ≥…‘±÷°“ΜΘ§ «Ρή”κBcl-2ΒΑΑΉΫαΚœΒΡΆ§‘¥Ζ÷Ή”Θ§ΝΫ’Ώ‘ΎœΗΑϊ÷–ΒΡ±»άΐΨωΕ®ΝΥœΗΑϊ «ΖώΒρΆωΓΘΒ±Bcl-2ΒΑΑΉ‘ΎœΗΑϊ÷–ΙΐΝΩ±μ¥ο ±Θ§”κΥυ”–BaxΫαΚœΘ§œΗΑϊ≥…ΜνΘ§ΒρΆω“÷÷ΤΘΜΒ±BaxΙΐΝΩ¥φ‘ΎΘ§”κBcl-2–Έ≥…“λ‘¥ΕΰΨέΧεΘ§‘ρœΗΑϊΒρΆω[13]ΓΘΥϋΟ«”κœΗΑϊΒρΆωΒΡΙΊœΒ‘Ύ÷ΉΝωΓΔΝήΑΆœΗΑϊΖΫΟφ“―”–¥σΝΩ―–ΨΩΘ§ΒΪ”κ…ζ÷≥œΗΑϊΒρΆωΒΡΙΊœΒ»¥Κή…Ό±®ΒάΓΘSuzukiΒ»[4]“‘«–≥ΐ–‘œΌΒΡ–έ–‘–Γ σ…ζ÷≥ΙήΒά(«ΑΝ–œΌΓΔΗΫΊΚ)ΈΣΒρΆωΡΘ–ΆΘ§ΖΔœ÷ΥφΉ≈ΒρΆωΒΡ≤ζ…ζ≥ωœ÷Bcl-2œ¬ΒςΘ§≤Δ‘Λ ΨΉ≈fasΜυ“ρΒΡ±μ¥οΓΘΕχΈ“Ο«ΒΡ―–ΨΩ‘ρΖΔœ÷Θ§‘Ύ’ΐ≥Θ–Γ σΊΚΆη…ζΨΪœΗΑϊΒΡΑϊ÷ ÷–”–¥σΝΩBcl-2ΒΑΑΉΓΔ…ΌΝΩBaxΒΑΑΉ±μ¥οΘ§“ΰΊΚ θΚσΒρΆωœΗΑϊΒΡ‘ωΦ”Θ§ΑιΥφΉ≈Bcl-2ΒΑΑΉΒΡΟςœ‘Φθ…ΌΘ§BaxΒΑΑΉΒΡΟςœ‘‘ωΕύΓΘΫαΙϊΧα Ψ‘ΎΊΚΆη…ζΨΪœΗΑϊΒΡΒρΆω÷–Θ§Bcl-2ΚΆBaxΜυ“ρ”–÷Ί“ΣΒςΫΎΉς”ΟΓΘ
ΊΚΆη…ζ÷≥œΗΑϊΒρΆωΦΑœύΙΊΜυ“ρΒςΫΎΉς”ΟΒΡ―–ΨΩΘ§ΈΣΡ––‘…ζ÷≥ΜζάμΒΡΧΫΧ÷ΧαΙ©ΝΥ“ΜΧθ–¬ΆΨΨΕΓΔΕ‘ΫΎ”ΐΓΔ≤Μ”ΐΒ»ΒΡ―–ΨΩΨυ”–÷Ί“Σ“β“εΓΘ
, ΑΌΡ¥“Ϋ“©
ΆΦ3 AΘ§B TUNEL»Ψ…ΪΓΝ300 A.’ΐ≥ΘΊΚΆη«–Τ§ B.“ΰΊΚ θΚσ8dΒΡΊΚΆη«–Τ§ ΦΐΆΖΥυ ΨΈΣ―τ–‘ΕύΚΥΨόœΗΑϊΓΘ(CΓΔDΓΔEΓΔF)…ζΈοΥΊ-ΩΙ…ζΈοΥΊΒΑΑΉDCSΦδΫ”Οβ“Ώ”ΪΙβ»Ψ…Ϊ ΓΝ300
C.Bcl-2ΒΑΑΉ‘Ύ’ΐ≥ΘΊΚΆη÷–ΒΡΖ÷≤Φ D.Bcl-2ΒΑΑΉ‘Ύ“ΰΊΚ θΚσ8dΒΡΊΚΆη÷–ΒΡΖ÷≤Φ E.BaxΒΑΑΉ‘Ύ’ΐ≥ΘΊΚΆη÷–ΒΡΖ÷≤Φ F.BaxΒΑΑΉ‘Ύ“ΰΊΚ θΚσ8dΒΡΊΚΆη÷–ΒΡΖ÷≤Φ Ήσœ¬Ϋ«ΈΣ“θ–‘Ε‘’’
Fig.3 A,B TUNEL staining of the formalin-fixed, paraffin-embedded testicular tissue sections. Postive staining appears green fluorescence. ΓΝ300 A.normal testes. B. cryptorchid testes on day 8. Arrow indicates positive Multinucleated giant cell.
, http://www.100md.com
(C,D,E,F)Biotin-avidin DCS system indirect immunofluorescence staining. Postive staining appears green fluorescence. ΓΝ300 C: Distribution of Bcl-2 protein in normal testes.D: Distribution of Bcl-2 protein in cryptorchid testes on day 8. E: Distribution of Bax protein in normal testes. F: Distribution of Bax protein in cryptorchid testes on day 8. Insert indicates negative control.
* ΙζΦ“Ή‘»ΜΩΤ―ßΜυΫπΉ ÷ζΩΈΧβ(No.39270380)ΚΆΈά…ζ≤ΩΩΤ―–ΜυΫπΉ ÷ζΩΈΧβ(No.94-2-004)
, ΑΌΡ¥“Ϋ“©
Γς Department of Histology and Embryology, Institute of Basic Medical Sciences, CAMS and PUMC, Beijing 10005, China
≤ΈΩΦΈΡœΉ
[1]Allan DJ, Harmon BV, Roberts SA. Spermatogonial apoptosis has three morphologically recognizable phases and shows no circadian rhythm during normal spermatogenesis in the rat. Cell Prolif, 1992,25(3):241
[2]Rodriguez JB, Garcia CM. Spontaneous germ cell death in the testis of the adult rat takes the form of apoptosis: re-evaluation of cell types that exhibit the ability to die during spermatogenesis. Cell Prolif, 1996,29(1):13
, ΑΌΡ¥“Ϋ“©
[3]Ν÷»ξœ…Ήέ ωΘ§Υο÷ΨœΆ…σ–Θ.bcl-2Μυ“ρΦ“Ήε”κœΗΑϊ≥Χ–ρ–‘ΥάΆω.ΙζΆβ“Ϋ―ß“≈¥Ϊ―ßΖ÷≤αΘ§1997Θ§20(5)ΘΚ242
[4]Suzuki A, Matsuzawa A, Iguchi T. Down regulation of bcl-2 is the first step on fas mediated apoptosis of male reproductive tract. Oncogene 1996,13(1):31
[5]Yin YZ, Hawkins KL, Dewolf WC, et al. Heat stress causes testicular germ cell apoptosis in adult mice. J Andro, 1997,18(2),159
[6]Xu ZL, Parker SB, Minkoff R. Distribution of type IV collagen, laminin, and fibronection during maxillary process formation in the chick embryo. Am J Anat, 1990,187(3):232
, ΑΌΡ¥“Ϋ“©
[7]Tapanainen JS, Tilly JL, Vihko KK, et al. Hormonal control of apoptotic cell death in the testis: Gonadotropins and androgens as testicular cell survival factors. Mol Endocrinol, 1993,7(5):643
[8]Shetty J,Marathe GK, Dighe RR, et al. Specific immunoneutralization of FSH leads to apoptotic cell death of the pachytene spermatocytes and spermatogonial cells in the rat. Endocrinol, 1996,137(5):2179
[9]Shikone T, Billig H, Hsueh AJW, et al. Experimentally induced cryptorchidism increases apoptosis in rat testis. Biolo Reproduc, 1994,51(5):865
, http://www.100md.com
[10]Heiskanen P, Billig H, Toppari J, et al. Apoptotic cell death in the normal and cryptorchid human testis: The effect of human chorionic gonadotropin on testicular cell survival. Pediatric Res, 1996,40(2):351
[11]Sharpe RM, Kerr JB, Fraser HM, et al. Intratesticular factors and testosterone secretion: effect of treatments that alter the level of testosterone within the testis. J Androl, 1986, 7(3):180
[12]Nguyen M, Branton PE, Walton PA, et al. Role of membrane anchor domain of bcl-2 in suppression of apoptosis caused by E1B-defective adenovirus. J Biol Chem, 1994,269(24):16521
[13]Oltrai ZN, Milliman CL, Korsmeyer SJ, et al. Bcl-2 heterodimerizes in vivo with a conserved homolog, Bax, that accelerates programmed cell death. Cell, 1993,74(4):609
’Ηε1998-01 –όΜΊ1998-05, ΑΌΡ¥“Ϋ“©