��Դ�Ծ����д������Ե�ˮ����Χ������ͻ���صı���
���ߣ����� ��¹ϣ
��λ������ ��¹ϣ(����ҽ�ƴ�ѧ����ѧϵ������ 100083)
�ؼ��ʣ�ͻ���أ���ˮ����Χ���ʣ���Դ�Ծ��ʣ���ȼ������
����ѧ��990306
��ժҪ�� Ŀ�� ������������ȼ����Դ�Ծ����д������Ե�ˮ����Χ����(PAG)��ͻ���ܶȵı仯��������� ������ϸ����ѧ�����ͼ����ͼ����������� ��� �����͵�ȼ����Դ�Ծ����д���PAG��ͻ����P38���߷�Ӧ�����У�����ܶ�ֵ(COD)��������������Wistar����ȼ����Դ�Ծ����д���PAG���༰���������������ͻ����P38���߷�Ӧ�����CODֵ��������������Դ�Ծ����д��� ���� PAG��ͻ���ܶȵĸı�����Դ�Ծ��ʼ����ȼ����������ء�
EXPRESSION OF SYNAPTOPHYSIN IN PERIAQUEDUCTAL GRAY
, http://www.100md.com
IN AUDIOGENIC SEIZURES-PRONE RATS
Liu Meng, Xu Luxi��
(Department of Anatomy, Beijing Medical University, Beijing)
��Abstract�� Objective To find out the changes of synaptic density in the periaqueductalgray(PAG) of the naive and kindled audiogenic seizures-prone rats. Methods Immunocytochemistry and computer image analysis technique were used.Results The value of corrected optical density (COD) of the synaptophysin P38 immunoreactivity in the PAG of the naive and kindled audiogenic seizures-prone rats were significantly higher than that of the normal Wistar rats. The COD value in the dorsal and dorsolateral subdivisions of the PAG in the kindled audiogenic seizures-prone rats were significantly higher than that in the naive audiogenc seizures-prone rats. Conclusion Changes of synaptic density in the PAG may be related to the susceptibility of audiogenic seizures and its kindling process.
, ��Ĵҽҩ
��Key words�� Synaptophysin; Periaqueductal gray; Audiogenic seizures; Kindling; Rat
ͻ����(synaptophysin)�ֳ�P38����һ�ַ�����Ϊ38kD�����Ըƽ���ǵ��ף�����Ϊһ������Ĥ���ף�������������������ĩ��ͻ��ǰ�ɷ��ڣ���ѡ���Եطֲ���Сͻ������Ĥ�İ�����[1]��ͻ����P38����Ca2+-�����Ե������ͷŹ��̣��������������ݵ���ͬ�����ʱ�̼���ʱ��ͻ�������Եĵ���[2]��ͬʱ��P38��Ϊͻ�����ݵ�һ�������Ա�־���ף��������ͷֲ��ܶȿɼ�ӷ�ӳͻ�����ܶ�[3]���ݴˣ���ʵ����P38����ϸ����ѧ��Ӧ��ǿ��Ϊָ�꣬��ӵض���Դ�Ծ����д���(P77PMC)���Ե�ˮ����Χ����(periaqueductal gray, PAG)������ͻ���ķֲ�����������о����ù⾵�۲�PAG���ڶθ�������Ƭ��ͻ���ܶȵı仯�������ڹ⾵ˮƽ���˽�PAG�ھ���״̬�³��ṹ��ijЩ�仯��
, ��Ĵҽҩ
���Ϻͷ���
1. ������Դ�Ծ��ʵ�ȼģ��
������Դ�Ծ����д���(P77PMC��ѡ��WistarƷϵ���ɱ���ҽ�ƴ�ѧʵ�鶯����������)��ÿ�����1�������̼�(106dB,60s)���շ�P77PMC����ǿֱ-��������Դ�Ծ��ʷ����������̼�40d��ɸѡ���о��ʷ�������20������Ϊ��ȼ����
2. ������鼰��Ƭ�Ʊ�
ʵ�鶯���Ϊ�������ԣ�����350��450g������3�飬��NW��(normal Wistar rats,n=4)������Wistar����NP��(naive P77PMC rats,n=4)��δ�ܴ�������Դ�Ծ����д���KP��(kindled P77PMC rats,n=4)����Դ�Ծ��ʵ�ȼ���P77PMC����
3�鶯���Ը���������(chloropent, 3 ml/kg)��ǻע��������������-����������ܣ������¹�ע������ˮ100��150 ml������ע��4%��ۼ�ȩ��0.1 mol/L�����λ���Һ(pH7.4��4��)300ml������֯����ͬ�̶�Һ�к�̶�24h��4�档������ˮ������ʯ��(54��56��)�����������϶�����������10 ��m��Ĺ�״������Ƭ������Ϳ�и�����������Ƭ�ϡ�
, ��Ĵҽҩ
3. P38����ϸ����ѧ����
3����Ƭ�����ױ�������0.01 mol/L������������ˮ����Һ(PBS��pH7.4)Ưϴ�������·ֱ��뺬0.3% Triton X-100��0.01 mol/L PBS����0.3% H2O2�Ĵ��״�(�����Դ�Թ�������ø)��2%������Ѫ��(���Ʒ�������IgG�Ľ��)��30 min��֮����С��P38����¡����(1��200��Sigma)������1h��֮����4����䣬24h���������������ؽ�ϵ�����С��IgG(1��200,Vector)���Ѱ���-������-��������ø������(ABC)1��100��Vector)�и�����1.5h�����¡����ϸ�����֮�����0.01 mol/L PBS���Ưϴ������Ժ�0.01% H2O2��0.04%DAB(��������������TCI)��Һ��ɫ5��10 min��������ˮ��������������Ƭ�����Զ�����Ƭ��0.01 mol/L PBS����С��P38����¡���壬���ಽ����ͬ��
, http://www.100md.com
4. �۲������
4.1 ���Թ۲죺������Ұ�����¹۲�P38���߷�Ӧ�������̬�ص㼰����PAG�������ڵķֲ��ص㡣
4.2 P38���߷�Ӧ������PAG�������ֲ��Ķ���������ͻ����P38���߷�Ӧ����ı��ǿ���Թ��ܶ�(optic density��A)ֵ����ʾ��ÿֻ�����PAG��״������Ƭ��ÿ��150 ��mȡƬ����ȡԼ15����Ƭ(�����ײ��2�ţ�β���3�ţ��ж�8��)���Լ����ͼ�����ϵͳ(Leica, Q550IW)����ÿ����Ƭ��������PAG��������P38���߷�Ӧ����Ĺ��ܶ�ֵ�������ڲ��������ϡ��������ϼ�������ɫ����ÿ����Ƭ�Դ˴��Ĺ��ܶ�ֵ��Ϊ����ֵ�����������ܶ�ֵ��ȥ����ֵ���õ�У�����ܶ�ֵ(corrected optic density,CA),����ͳ�Ʒ���ʱ���ô�У�����ܶ�ֵ������������������ɫ�Խ����Ӱ�졣����ÿ�����PAG��ÿ���������õ�120��CAֵ�������CAֵ��SPSS for Windowsͳ�Ʒ����������е����صķ������(one-way ANOVA)��
, http://www.100md.com
���� �������PAG��P38���߷�Ӧ�����CAֵ(��100)
Table CA value of P38 immunoreactivity in PAG in rats of groups(��100)
NW
NP
KP
ANOVA��
NW
NP
KP
�ڲ�����
medial subdivision
, ��Ĵҽҩ
2.06��0.99
2.87��0.89
3.50��0.73
A
B
B
���������
ventrolateral subdvision
1.75��0.62
3.17��1.19
4.00��1.35
A
, http://www.100md.com
B
B
��������
dorsal subdivision
1.92��0.67
3.17��0.94
5.58��1.73
A
B
C
���������
dorsolateral subdivision
, http://www.100md.com
1.38��0.74
2.90��0.75
4.88��1.81
A
B
C
*ANOVAΪ���������A��B��C����ͬ��ĸ֮���ʾ�������Բ���(P<0.05)����ͬ��ĸ��ʾ�������Բ���(P>0.05)
*ANOVA:analysis of variance. A B C: There is significant difference between different letters (P<0.05) and no significant difference between same letters(P>0.05)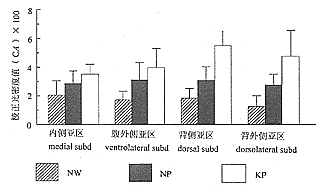
, ��Ĵҽҩ
ͼ5 �������PAG��P38���߷�Ӧ�����У�����ܶ�ֵ(��100)
Fig. 5 CA value of P38 immunoreactivity in PAG in rats of groups(��100)
�� ��
P38���߷�Ӧ����Ϊ�ػ�ɫ��״�����״��������Ҫ�ֲ�����ձ�ڣ���Щ����Χ������Ԫ��Χ�����г���Ԫ����������������Ĵ�С��һ������ͻ��ǰ��ĩ�Ĵ�С��ͻ�������йأ������ڿ���״������ټ������ʡ�Ѫ�ܼ�����ϸ������ɫ�����Զ�����Ƭδ��P38���߷�Ӧ���
�����������������PAG��P38���߷�Ӧ����ȽϷḻ����ɫ����������Χ���ϡ������Dz������ṹ����PAG�����������⾵�¿ɼ�ÿһ�����ڲ�P38���߷�Ӧ����ķֲ�Ҳ��Ϊ���ȣ�������֮��Ⱦɫ��dz�IJ��쾵�²������𣻸������֮�䣬�ɼ�PAG��ɫ��dz�����Բ���(ͼ1��4)��CAֵ�Ķ���������������ͼ5��
, ��Ĵҽҩ
�� ��
1. ͻ����P38�����ʼ�������
ͻ����P38��һ�ַ�����Ϊ38kD�����Ըƽ���ǵ��ף������Եطֲ��ڼ�������������ĩ��СԲ�λ����(����)ͻ��ǰ����Ĥ����Ĥ�棬��һ����Ҫ������Ĥ��־����[1]���⾵���������Ŀ���״���Բ���ʵ���Ͼ��dzɴ����е�ͻ������[4]������P38�ĺ�����ͻ���ܶ�����أ����ͨ������P38�������Բ����ƽ�����ܶ�ֵ�����Է�ӳ��ͻ��ǰ���ݵķֲ�����������Ʋ�ͻ���������ܶȵı仯���Ӷ��ﵽ�ڹ⾵ˮƽ�������ṹ��Ŀ�ġ��������������о���P38Ϊָ����̽��Alzheimer����Huntington������ϵͳ������˥��״̬��ͻ���������ܶȵı仯��������P38��˥����ͻ����ʧ�������[5��6]��
���������о�������P38��Ϊһ����������Ĥ���ף����������ͻ�������йصĻ��������Ca2+-�����Ե������ͷŹ��̣����볤ʱ��ͻ��������������ء���ʱ����ǿ(long-term potentiation, LTP)���յ���ά�ְ�����ͻ����̬�Ŀ����Ա仯����̬�Ŀ����Ըı�������ʹͻ���Ĺ�����ǿ��������̬�ϵı仯����ͻ��ǰ�ı��ͻ����ı䣬�����������Ӽ��ֲ��ı仯��ͻ��ǰ�ı���Ҫ��ʽ�������������������ֱ�Ȼ������P38�ϳɵ����ӡ���һ���棬��˥�ϼ�������������£�LPT�������½���P38�ĺ�������������Ҳ����[7��8]��
, ��Ĵҽҩ
��ȼ���յ���Ԫ��ͻ������ͻ���ؽ�����ͻ���γɵȿ����Ա仯[9��10]��ͬʱ����ȼ������Ҫ�е��ĺϳ�[11]���ɴ˿��Ʋ��ȼ����ҲӦ��P38�ĺϳɡ��ɼ���P38��������ϵͳ�����Ե���Чָ��֮һ��
2. PAG�ڵ�ͻ����P38����Դ�Ծ��ʼ����ȼ�Ĺ�ϵ
��ʵ��Ľ����ʾ��(1) NP�����PAG��������P38�����NW��������ǿ��˵��P77PMC����PAG������Wistar����PAG֮���Ѿ��ڳ��ṹ�ϣ���ͻ��ˮƽ�ϳ����˲��죬P77PMC����PAG�ڵ�ͻ���ܶȻ������ܶȽ�����Wistar�����������ߣ�����P77PMC������ɸѡ�����Ѵ���23��������ͻ���ܶȵı仯����Դ��P77PMC����ij���Ŵ����صĸı䣬�˸ı䵼����PAG����Ԫ�����ֻ����쳣���Ӷ�ʹ��ͻ���ṹ�����ı䣬�Ʋ�����ͻ��ˮƽ�ı仯������P77PMC����ľ��������йأ���������в������(2) ������Դ�Ծ��ʷ������µ�ȼ��PAG���༰���������P38����ˮƽ��NW��NP����������ǿ��˵����Դ�Ծ��ʷ���������ʹP77PMC����PAG�ڷ������ԵĿ����Ըı䣬ͻ���ܶȻ������ܶ��������ӡ����ֿ����Ա仯�Ļ��ƿ��������������c-fos����ķ��������йأ�c-fos��Ϊ���ڵ�����ʹ�����������͵��ذл���ı����ijЩ�л���ı��������Գ�ʱ�̵�����Ԫ�Ĺ��ܣ�һ���Ե�c-fos������ܲ���������л����������Ըı䣬���������ʷ������µ�PAG��������������ڴ�����Ԫ����c-fos����������ջ�ʹijЩ��������صİл���ı���������ǿ���Ӷ�ʹ��ת¼���ϳɵIJ������ͻ���������ṹ�ȷ���Ŀ����Ըı䡣��֪����ϸ���Ǽܵ��ס�ͻ�����ݵ��ס�����ͨ��������Ƚṹ���Ļ��������ܼ������ڻ�����صİл���[12]����ĿǰΪֹ����c-fos���������Щ�л�����Щ�л��������������ϸ���ķ�Ӧ�Լ���Щ��Ӧ������Ӱ����֪֮���٣����ҹ��ڵ�ȼ��������̬ѧ�仯���о�Ҳ����һ�½����
, ��Ĵҽҩ
��һ�ֿ����Ա仯�Ŀ��ܻ�����LTP���ƣ��������ʷ�������PAG����༰���������ڴ����ĹȰ�������Ԫ�˷ܣ��Ȱ����ͷų������࣬ͨ��Ca2+��NMDA���弰��л�ԹȰ������塢������ʹ(һ������)��һϵ�����ʵIJ��룬����ʹͻ����̬�����仯��ͻ����̬�仯�ֵ���ͻ�����ܵ���ǿ[7]������ȼ��LTP��ȷ����ijЩ��ͬ�����ƣ����߿����[13��14]��
PAG������������ر���β��εĸ���������ڹȰ�������Ԫ���ٶ�GABA����Ԫ�϶࣬��һ����Ҫ�������������õ����࣬�����ױ�������ȼ������Ҫ�������˷��Ե���ǿ�����������Եļ�����ʵ�ֵ�[15]���������ܲ��μ�ͻ�������Եı仯������Fos���İл�����ܲ��Ǻϳ�ͻ�����ݵ��Ļ���P38����û�����Ե���ǿ��ͬ�������ڴ�����ȱ���ḻ���˷������ἰ�����壬��LTP�������Ա仯Ҳ�����ԡ�
��ʵ����Ȼ��ʾ����������ȼ��P77PMC����PAG��ͻ���ܶȵı仯����������ͻ�����͡�ͻ�����ʡ�����ͻ��(perforated synapse)���ʡ����Դ����ȵȷ�ӳͻ�������Ե�ָ��ĸı�����羵�о�֤ʵ��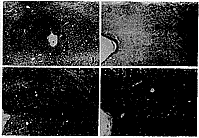
, http://www.100md.com
ͼ �� ˵ ��
ͼ1 KP�����PAG��P38���߷�Ӧ����ķֲ� ��70
ͼ2��4 NW��(ͼ2)��NP��(ͼ3)��KP��(ͼ4)����PAG��P38���߷�Ӧ����ķֲ� ��150 D.�������� DL.���������
Explanation of figures
Fig.1 The distribution of P38 immunoreactivity in PAG of KP rats ��70
Fig.2-4 The distribution of P38 immunoreactivity in PAG of NW(Fig.2), NP(Fig.3) and KP (Fig.4) rats ��110 D��dorsal subdivision DL��dorsolateral subdivision
, http://www.100md.com
�� Department of Anatomy, Beijing Medical University, Beijing 100083,China
�����
[1]Wiedenmann B, Franke W. Identification and localization of synaptophysin, an integral membrane glycoprotein of Mr 38 000 characteristic of presynaptic vesicles. Cell,1985,41(3)��1017
[2]Catsicas S, Grenningloh G, Pich EM. Nerve-terminal proteins: to fuse to learn. TINS, 1994,17(9):368
, ��Ĵҽҩ
[3]Saito S, Kobayashi S, Ohashi T. Decreased synaptic density in aged brains and its prevention by rearing under enriched enviroment as revealed by synaptophysin contents. J Neuroscience Reasearch,1994,39(1):157
[4]Masliah E, Terry RD, Alford M, et al. Quantitative immunohistochemistry of synaptophysin in human neocortex:an alternative method to estimate density of presynaptic terminal in paraffin sections. J Histochem and Cytochem,1990,38(6):837
, ��Ĵҽҩ
[5]Masliah E, Terry RD, Alford M, et al. Cortical and subcortical patterns of synaptophysin-like immunoreactivity in Alzheimer's disease. Am J Pathol, 1991,138(1):235
[6]Goto S, Hirano A. Synaptophysin expression in the striatum in Huntington's disease. Aca Neuropathol,1993,80(1):88
[7]Lynch MA, Voss KL, Rodriguez J, et al. Increase in synaptic vesicle proteins accompanies long-term potentiation in the dentate gyrus. Neurosci,1994��60(1):1
, http://www.100md.com
[8]Lynch MA, Mullany P. Changes in protein synthesis and synthesis of the synaptic vesicle protein, synaptophysin, in entorhinal cortex following induction of long-term potentiation in dentate gyrus:an age-related study in the rat. Neuropharmacology,1997,36(7):973
[9]Hexx ST, Cavazos J. Synaptic recognization in the hippocampus induced by abnormal functional activity. Science, 1988,239(4844):1147
[10]Chang FL, Hawrylak N, Greenough WT. Astrocytic and synaptic response to kindling in hippocampal subfield CA1. I synaptogenesis in response to kindling in vitro. Brain Res,1993,603(2):309
, ��Ĵҽҩ
[11]Cain D, Corcoran M, Staines W. Effects of protein. synthesis inhibition on kindling in the mouse. Exp Neurol.1980��68(3):409
[12]Morgan JI, Curran T. Proto-oncogene transcription factors and epilepsy. TINS, 1991,12(9):343
[13]Majkowski J. Kindling:a model for epilepsy and memory. Acta Neurol Scand, 1986,74(suppl 109):97
[14]Cain DP. Long-term potentiation and kindling:how similar are the mechanisms? TINS, 1989,12(1):6
[15]Cottrell GA, Robertson HA. Baclonfen exacerbates epileptic myoclonus in kindled rats. Neuropharmacology,1987,26(6):645
�ո�1998-12 ��1998-03, ��Ĵҽҩ
��λ������ ��¹ϣ(����ҽ�ƴ�ѧ����ѧϵ������ 100083)
�ؼ��ʣ�ͻ���أ���ˮ����Χ���ʣ���Դ�Ծ��ʣ���ȼ������
����ѧ��990306
��ժҪ�� Ŀ�� ������������ȼ����Դ�Ծ����д������Ե�ˮ����Χ����(PAG)��ͻ���ܶȵı仯��������� ������ϸ����ѧ�����ͼ����ͼ����������� ��� �����͵�ȼ����Դ�Ծ����д���PAG��ͻ����P38���߷�Ӧ�����У�����ܶ�ֵ(COD)��������������Wistar����ȼ����Դ�Ծ����д���PAG���༰���������������ͻ����P38���߷�Ӧ�����CODֵ��������������Դ�Ծ����д��� ���� PAG��ͻ���ܶȵĸı�����Դ�Ծ��ʼ����ȼ����������ء�
EXPRESSION OF SYNAPTOPHYSIN IN PERIAQUEDUCTAL GRAY
, http://www.100md.com
IN AUDIOGENIC SEIZURES-PRONE RATS
Liu Meng, Xu Luxi��
(Department of Anatomy, Beijing Medical University, Beijing)
��Abstract�� Objective To find out the changes of synaptic density in the periaqueductalgray(PAG) of the naive and kindled audiogenic seizures-prone rats. Methods Immunocytochemistry and computer image analysis technique were used.Results The value of corrected optical density (COD) of the synaptophysin P38 immunoreactivity in the PAG of the naive and kindled audiogenic seizures-prone rats were significantly higher than that of the normal Wistar rats. The COD value in the dorsal and dorsolateral subdivisions of the PAG in the kindled audiogenic seizures-prone rats were significantly higher than that in the naive audiogenc seizures-prone rats. Conclusion Changes of synaptic density in the PAG may be related to the susceptibility of audiogenic seizures and its kindling process.
, ��Ĵҽҩ
��Key words�� Synaptophysin; Periaqueductal gray; Audiogenic seizures; Kindling; Rat
ͻ����(synaptophysin)�ֳ�P38����һ�ַ�����Ϊ38kD�����Ըƽ���ǵ��ף�����Ϊһ������Ĥ���ף�������������������ĩ��ͻ��ǰ�ɷ��ڣ���ѡ���Եطֲ���Сͻ������Ĥ�İ�����[1]��ͻ����P38����Ca2+-�����Ե������ͷŹ��̣��������������ݵ���ͬ�����ʱ�̼���ʱ��ͻ�������Եĵ���[2]��ͬʱ��P38��Ϊͻ�����ݵ�һ�������Ա�־���ף��������ͷֲ��ܶȿɼ�ӷ�ӳͻ�����ܶ�[3]���ݴˣ���ʵ����P38����ϸ����ѧ��Ӧ��ǿ��Ϊָ�꣬��ӵض���Դ�Ծ����д���(P77PMC)���Ե�ˮ����Χ����(periaqueductal gray, PAG)������ͻ���ķֲ�����������о����ù⾵�۲�PAG���ڶθ�������Ƭ��ͻ���ܶȵı仯�������ڹ⾵ˮƽ���˽�PAG�ھ���״̬�³��ṹ��ijЩ�仯��
, ��Ĵҽҩ
���Ϻͷ���
1. ������Դ�Ծ��ʵ�ȼģ��
������Դ�Ծ����д���(P77PMC��ѡ��WistarƷϵ���ɱ���ҽ�ƴ�ѧʵ�鶯����������)��ÿ�����1�������̼�(106dB,60s)���շ�P77PMC����ǿֱ-��������Դ�Ծ��ʷ����������̼�40d��ɸѡ���о��ʷ�������20������Ϊ��ȼ����
2. ������鼰��Ƭ�Ʊ�
ʵ�鶯���Ϊ�������ԣ�����350��450g������3�飬��NW��(normal Wistar rats,n=4)������Wistar����NP��(naive P77PMC rats,n=4)��δ�ܴ�������Դ�Ծ����д���KP��(kindled P77PMC rats,n=4)����Դ�Ծ��ʵ�ȼ���P77PMC����
3�鶯���Ը���������(chloropent, 3 ml/kg)��ǻע��������������-����������ܣ������¹�ע������ˮ100��150 ml������ע��4%��ۼ�ȩ��0.1 mol/L�����λ���Һ(pH7.4��4��)300ml������֯����ͬ�̶�Һ�к�̶�24h��4�档������ˮ������ʯ��(54��56��)�����������϶�����������10 ��m��Ĺ�״������Ƭ������Ϳ�и�����������Ƭ�ϡ�
, ��Ĵҽҩ
3. P38����ϸ����ѧ����
3����Ƭ�����ױ�������0.01 mol/L������������ˮ����Һ(PBS��pH7.4)Ưϴ�������·ֱ��뺬0.3% Triton X-100��0.01 mol/L PBS����0.3% H2O2�Ĵ��״�(�����Դ�Թ�������ø)��2%������Ѫ��(���Ʒ�������IgG�Ľ��)��30 min��֮����С��P38����¡����(1��200��Sigma)������1h��֮����4����䣬24h���������������ؽ�ϵ�����С��IgG(1��200,Vector)���Ѱ���-������-��������ø������(ABC)1��100��Vector)�и�����1.5h�����¡����ϸ�����֮�����0.01 mol/L PBS���Ưϴ������Ժ�0.01% H2O2��0.04%DAB(��������������TCI)��Һ��ɫ5��10 min��������ˮ��������������Ƭ�����Զ�����Ƭ��0.01 mol/L PBS����С��P38����¡���壬���ಽ����ͬ��
, http://www.100md.com
4. �۲������
4.1 ���Թ۲죺������Ұ�����¹۲�P38���߷�Ӧ�������̬�ص㼰����PAG�������ڵķֲ��ص㡣
4.2 P38���߷�Ӧ������PAG�������ֲ��Ķ���������ͻ����P38���߷�Ӧ����ı��ǿ���Թ��ܶ�(optic density��A)ֵ����ʾ��ÿֻ�����PAG��״������Ƭ��ÿ��150 ��mȡƬ����ȡԼ15����Ƭ(�����ײ��2�ţ�β���3�ţ��ж�8��)���Լ����ͼ�����ϵͳ(Leica, Q550IW)����ÿ����Ƭ��������PAG��������P38���߷�Ӧ����Ĺ��ܶ�ֵ�������ڲ��������ϡ��������ϼ�������ɫ����ÿ����Ƭ�Դ˴��Ĺ��ܶ�ֵ��Ϊ����ֵ�����������ܶ�ֵ��ȥ����ֵ���õ�У�����ܶ�ֵ(corrected optic density,CA),����ͳ�Ʒ���ʱ���ô�У�����ܶ�ֵ������������������ɫ�Խ����Ӱ�졣����ÿ�����PAG��ÿ���������õ�120��CAֵ�������CAֵ��SPSS for Windowsͳ�Ʒ����������е����صķ������(one-way ANOVA)��
, http://www.100md.com
���� �������PAG��P38���߷�Ӧ�����CAֵ(��100)
Table CA value of P38 immunoreactivity in PAG in rats of groups(��100)
NW
NP
KP
ANOVA��
NW
NP
KP
�ڲ�����
medial subdivision
, ��Ĵҽҩ
2.06��0.99
2.87��0.89
3.50��0.73
A
B
B
���������
ventrolateral subdvision
1.75��0.62
3.17��1.19
4.00��1.35
A
, http://www.100md.com
B
B
��������
dorsal subdivision
1.92��0.67
3.17��0.94
5.58��1.73
A
B
C
���������
dorsolateral subdivision
, http://www.100md.com
1.38��0.74
2.90��0.75
4.88��1.81
A
B
C
*ANOVAΪ���������A��B��C����ͬ��ĸ֮���ʾ�������Բ���(P<0.05)����ͬ��ĸ��ʾ�������Բ���(P>0.05)
*ANOVA:analysis of variance. A B C: There is significant difference between different letters (P<0.05) and no significant difference between same letters(P>0.05)
, ��Ĵҽҩ
ͼ5 �������PAG��P38���߷�Ӧ�����У�����ܶ�ֵ(��100)
Fig. 5 CA value of P38 immunoreactivity in PAG in rats of groups(��100)
�� ��
P38���߷�Ӧ����Ϊ�ػ�ɫ��״�����״��������Ҫ�ֲ�����ձ�ڣ���Щ����Χ������Ԫ��Χ�����г���Ԫ����������������Ĵ�С��һ������ͻ��ǰ��ĩ�Ĵ�С��ͻ�������йأ������ڿ���״������ټ������ʡ�Ѫ�ܼ�����ϸ������ɫ�����Զ�����Ƭδ��P38���߷�Ӧ���
�����������������PAG��P38���߷�Ӧ����ȽϷḻ����ɫ����������Χ���ϡ������Dz������ṹ����PAG�����������⾵�¿ɼ�ÿһ�����ڲ�P38���߷�Ӧ����ķֲ�Ҳ��Ϊ���ȣ�������֮��Ⱦɫ��dz�IJ��쾵�²������𣻸������֮�䣬�ɼ�PAG��ɫ��dz�����Բ���(ͼ1��4)��CAֵ�Ķ���������������ͼ5��
, ��Ĵҽҩ
�� ��
1. ͻ����P38�����ʼ�������
ͻ����P38��һ�ַ�����Ϊ38kD�����Ըƽ���ǵ��ף������Եطֲ��ڼ�������������ĩ��СԲ�λ����(����)ͻ��ǰ����Ĥ����Ĥ�棬��һ����Ҫ������Ĥ��־����[1]���⾵���������Ŀ���״���Բ���ʵ���Ͼ��dzɴ����е�ͻ������[4]������P38�ĺ�����ͻ���ܶ�����أ����ͨ������P38�������Բ����ƽ�����ܶ�ֵ�����Է�ӳ��ͻ��ǰ���ݵķֲ�����������Ʋ�ͻ���������ܶȵı仯���Ӷ��ﵽ�ڹ⾵ˮƽ�������ṹ��Ŀ�ġ��������������о���P38Ϊָ����̽��Alzheimer����Huntington������ϵͳ������˥��״̬��ͻ���������ܶȵı仯��������P38��˥����ͻ����ʧ�������[5��6]��
���������о�������P38��Ϊһ����������Ĥ���ף����������ͻ�������йصĻ��������Ca2+-�����Ե������ͷŹ��̣����볤ʱ��ͻ��������������ء���ʱ����ǿ(long-term potentiation, LTP)���յ���ά�ְ�����ͻ����̬�Ŀ����Ա仯����̬�Ŀ����Ըı�������ʹͻ���Ĺ�����ǿ��������̬�ϵı仯����ͻ��ǰ�ı��ͻ����ı䣬�����������Ӽ��ֲ��ı仯��ͻ��ǰ�ı���Ҫ��ʽ�������������������ֱ�Ȼ������P38�ϳɵ����ӡ���һ���棬��˥�ϼ�������������£�LPT�������½���P38�ĺ�������������Ҳ����[7��8]��
, ��Ĵҽҩ
��ȼ���յ���Ԫ��ͻ������ͻ���ؽ�����ͻ���γɵȿ����Ա仯[9��10]��ͬʱ����ȼ������Ҫ�е��ĺϳ�[11]���ɴ˿��Ʋ��ȼ����ҲӦ��P38�ĺϳɡ��ɼ���P38��������ϵͳ�����Ե���Чָ��֮һ��
2. PAG�ڵ�ͻ����P38����Դ�Ծ��ʼ����ȼ�Ĺ�ϵ
��ʵ��Ľ����ʾ��(1) NP�����PAG��������P38�����NW��������ǿ��˵��P77PMC����PAG������Wistar����PAG֮���Ѿ��ڳ��ṹ�ϣ���ͻ��ˮƽ�ϳ����˲��죬P77PMC����PAG�ڵ�ͻ���ܶȻ������ܶȽ�����Wistar�����������ߣ�����P77PMC������ɸѡ�����Ѵ���23��������ͻ���ܶȵı仯����Դ��P77PMC����ij���Ŵ����صĸı䣬�˸ı䵼����PAG����Ԫ�����ֻ����쳣���Ӷ�ʹ��ͻ���ṹ�����ı䣬�Ʋ�����ͻ��ˮƽ�ı仯������P77PMC����ľ��������йأ���������в������(2) ������Դ�Ծ��ʷ������µ�ȼ��PAG���༰���������P38����ˮƽ��NW��NP����������ǿ��˵����Դ�Ծ��ʷ���������ʹP77PMC����PAG�ڷ������ԵĿ����Ըı䣬ͻ���ܶȻ������ܶ��������ӡ����ֿ����Ա仯�Ļ��ƿ��������������c-fos����ķ��������йأ�c-fos��Ϊ���ڵ�����ʹ�����������͵��ذл���ı����ijЩ�л���ı��������Գ�ʱ�̵�����Ԫ�Ĺ��ܣ�һ���Ե�c-fos������ܲ���������л����������Ըı䣬���������ʷ������µ�PAG��������������ڴ�����Ԫ����c-fos����������ջ�ʹijЩ��������صİл���ı���������ǿ���Ӷ�ʹ��ת¼���ϳɵIJ������ͻ���������ṹ�ȷ���Ŀ����Ըı䡣��֪����ϸ���Ǽܵ��ס�ͻ�����ݵ��ס�����ͨ��������Ƚṹ���Ļ��������ܼ������ڻ�����صİл���[12]����ĿǰΪֹ����c-fos���������Щ�л�����Щ�л��������������ϸ���ķ�Ӧ�Լ���Щ��Ӧ������Ӱ����֪֮���٣����ҹ��ڵ�ȼ��������̬ѧ�仯���о�Ҳ����һ�½����
, ��Ĵҽҩ
��һ�ֿ����Ա仯�Ŀ��ܻ�����LTP���ƣ��������ʷ�������PAG����༰���������ڴ����ĹȰ�������Ԫ�˷ܣ��Ȱ����ͷų������࣬ͨ��Ca2+��NMDA���弰��л�ԹȰ������塢������ʹ(һ������)��һϵ�����ʵIJ��룬����ʹͻ����̬�����仯��ͻ����̬�仯�ֵ���ͻ�����ܵ���ǿ[7]������ȼ��LTP��ȷ����ijЩ��ͬ�����ƣ����߿����[13��14]��
PAG������������ر���β��εĸ���������ڹȰ�������Ԫ���ٶ�GABA����Ԫ�϶࣬��һ����Ҫ�������������õ����࣬�����ױ�������ȼ������Ҫ�������˷��Ե���ǿ�����������Եļ�����ʵ�ֵ�[15]���������ܲ��μ�ͻ�������Եı仯������Fos���İл�����ܲ��Ǻϳ�ͻ�����ݵ��Ļ���P38����û�����Ե���ǿ��ͬ�������ڴ�����ȱ���ḻ���˷������ἰ�����壬��LTP�������Ա仯Ҳ�����ԡ�
��ʵ����Ȼ��ʾ����������ȼ��P77PMC����PAG��ͻ���ܶȵı仯����������ͻ�����͡�ͻ�����ʡ�����ͻ��(perforated synapse)���ʡ����Դ����ȵȷ�ӳͻ�������Ե�ָ��ĸı�����羵�о�֤ʵ��
, http://www.100md.com
ͼ �� ˵ ��
ͼ1 KP�����PAG��P38���߷�Ӧ����ķֲ� ��70
ͼ2��4 NW��(ͼ2)��NP��(ͼ3)��KP��(ͼ4)����PAG��P38���߷�Ӧ����ķֲ� ��150 D.�������� DL.���������
Explanation of figures
Fig.1 The distribution of P38 immunoreactivity in PAG of KP rats ��70
Fig.2-4 The distribution of P38 immunoreactivity in PAG of NW(Fig.2), NP(Fig.3) and KP (Fig.4) rats ��110 D��dorsal subdivision DL��dorsolateral subdivision
, http://www.100md.com
�� Department of Anatomy, Beijing Medical University, Beijing 100083,China
�����
[1]Wiedenmann B, Franke W. Identification and localization of synaptophysin, an integral membrane glycoprotein of Mr 38 000 characteristic of presynaptic vesicles. Cell,1985,41(3)��1017
[2]Catsicas S, Grenningloh G, Pich EM. Nerve-terminal proteins: to fuse to learn. TINS, 1994,17(9):368
, ��Ĵҽҩ
[3]Saito S, Kobayashi S, Ohashi T. Decreased synaptic density in aged brains and its prevention by rearing under enriched enviroment as revealed by synaptophysin contents. J Neuroscience Reasearch,1994,39(1):157
[4]Masliah E, Terry RD, Alford M, et al. Quantitative immunohistochemistry of synaptophysin in human neocortex:an alternative method to estimate density of presynaptic terminal in paraffin sections. J Histochem and Cytochem,1990,38(6):837
, ��Ĵҽҩ
[5]Masliah E, Terry RD, Alford M, et al. Cortical and subcortical patterns of synaptophysin-like immunoreactivity in Alzheimer's disease. Am J Pathol, 1991,138(1):235
[6]Goto S, Hirano A. Synaptophysin expression in the striatum in Huntington's disease. Aca Neuropathol,1993,80(1):88
[7]Lynch MA, Voss KL, Rodriguez J, et al. Increase in synaptic vesicle proteins accompanies long-term potentiation in the dentate gyrus. Neurosci,1994��60(1):1
, http://www.100md.com
[8]Lynch MA, Mullany P. Changes in protein synthesis and synthesis of the synaptic vesicle protein, synaptophysin, in entorhinal cortex following induction of long-term potentiation in dentate gyrus:an age-related study in the rat. Neuropharmacology,1997,36(7):973
[9]Hexx ST, Cavazos J. Synaptic recognization in the hippocampus induced by abnormal functional activity. Science, 1988,239(4844):1147
[10]Chang FL, Hawrylak N, Greenough WT. Astrocytic and synaptic response to kindling in hippocampal subfield CA1. I synaptogenesis in response to kindling in vitro. Brain Res,1993,603(2):309
, ��Ĵҽҩ
[11]Cain D, Corcoran M, Staines W. Effects of protein. synthesis inhibition on kindling in the mouse. Exp Neurol.1980��68(3):409
[12]Morgan JI, Curran T. Proto-oncogene transcription factors and epilepsy. TINS, 1991,12(9):343
[13]Majkowski J. Kindling:a model for epilepsy and memory. Acta Neurol Scand, 1986,74(suppl 109):97
[14]Cain DP. Long-term potentiation and kindling:how similar are the mechanisms? TINS, 1989,12(1):6
[15]Cottrell GA, Robertson HA. Baclonfen exacerbates epileptic myoclonus in kindled rats. Neuropharmacology,1987,26(6):645
�ո�1998-12 ��1998-03, ��Ĵҽҩ