CRE和AP-1元件参与人hsp90β基因的表达
作者:刘秉胜 吴宁华 沈珝琲
单位:刘秉胜(中国医学科学院基础医学研究所 中国协和医科大学基础医学院 医学分子生物学国家重点实验室,北京100005);吴宁华(中国医学科学院基础医学研究所 中国协和医科大学基础医学院 医学分子生物学国家重点实验室,北京100005);沈珝琲(中国医学科学院基础医学研究所 中国协和医科大学基础医学院 医学分子生物学国家重点实验室,北京100005)
关键词:CRE元件;AP-1元件;hsp90b基因;基因表达
基础医学与临床000306
摘要:为了进一步阐明人热休克蛋白90b(hsp90b)基因的表达调控机制,本文对hsp90b基因调控序列中的CRE和 AP-1元件进行了研究。构建了含有这两个元件的系列删切报告基因质粒,通过转染 Jurkat细胞,检测并分析报告基因的活性。结果表明,CRE和 AP-1元件均不同程度地参与了hsp90b基因的组成性、PHA激活和热诱导表达。
, http://www.100md.com
中图分类号:Q786 文献标识码:A 文章编号:1001-6352(2000)03-0028-04
CRE and AP-1 elements in the expression of human hsp90b gene
LIU Bing-sheng,WU Ning-hua,SHEN Yu-fei
(National Laboratory of Medical Molecular Biology,Institute of Basic Medical Sciences,Chinese Academy
of Medical Science&Peking Union Medical College,Beijing100005,China)
, 百拇医药 Abstract:Several reporter plasmids with deleting CRE and AP-1 elements in the regulation sequences of hsp90b gene were constructed and transfected into Jurkat cells.Then the reporter genes' activity was detected.The results show that CRE and AP-1 elements were critical to the high constitutive expression of hsp90b gene; in the PHA activated expression,CRE or AP-1 elements play synergistic role; CRE and AP-1 elements also partially mediate in the heat shock inducible expression of hsp90b gene.
, http://www.100md.com
Key words: CRE; AP-1 element; hsp90b gene; gene expression
hsp90基因是热休克蛋白基因家族的成员。与其它热休克蛋白基因(如hsp70)不同hsp90基因通常在细胞内就有较高水平的组成性表达,并可进一步被热和丝裂原诱导[1]。 hsp90基因有a和b两个拷贝[2,3]。本组对hsp90b基因的表达顺式调节机理进行了研究。研究表明,上游启动子元件、核心启动子和第一外显子或第一内含子对于高组成性表达起重要作用;在热诱导表达中,第一内含子中两个典型的 HSE起了非常重要的作用[4]。我们在分析这些区域的顺式元件分布时,发现在上游启动区-127/-120bp处为环腺苷酸反应元件 (cAMP responsive element,简称 CRE)或称为转录激活因子结合位点 (the activating transcription factor,简称 ATF),同时在第一内含子+484/+489bp处有一 AP-1(the activator protein l)元件(或TRE元件, the phorbol 12-0-tetradecanoate 13-acetate responsive element,简称TRE)。 CRE元件的序列是:TGACGTCA。AP-1元件的序列是:TGACTCA。这两种元件可分别被CRE结合蛋白 CREB家族成员和 AP-1识别蛋白Jun/Fos所识别。研究表明这些转录因子均与信号传导有关[5~7]。因此,为了进一步深入了解 hsp90b基因的表达调控机理,我们对这两个元件在hsp90b基因表达调控中的作用进行了研究。
, 百拇医药
1 材料与方法
1.1 材料
1.1.1 主要试剂:Hinc、XhoI、BamHI、SmaI等限制性内切酶和T4 DNA连接酶为Boehinger Mannheim公司产品;其它限制性内切酶及DNA分子量标准均为协和友谊公司产品;PHA为Sigma公司产品;细胞裂解及荧光素酶检测试剂盒为Promega公司产品;3H-乙酰辅酶A为Amersham公司产品;BCA 蛋白分析试剂为Pierce公司产品。
1.1.2 质粒与细胞株:含有hsp90b基因调控序列-1039至+1531bp的CAT基因报告质粒b3.1和荧光素酶基因报告质粒b1.11由本课题组提供;Jurkat细胞为本课题组保存的细胞株。
1.2 方法
1.2.1 CRE和AP-1元件系缺失质粒的构建:b1.14经BamHI和XhoI部分酶解所得片段-173/+1351插入PMC/Luc报告质粒而建成;b1.15:由b1.11经Hinc和XhoI部分酶切所得片段-91/+1531插入经SmaI和XhoI酶切的PMC/Luc报告质粒建成;b1.14':由b1.14经SmaI和MluI酶解所得大片段用Klenow酶补平后自连而成;b1.15'由b1.15经SmaI和MluI酶解所得大片段用Klenow补平后自连而成。b3.2:b3.1经BamHI和XhoI部分酶解所得片段-173/+1531插入被SmaI和XhoI酶解的pBLCAT3报告质粒而建成;b3.4:b3.1经PstI完全酶解,Klenow酶补平后经Hinc部分酶解所得大片段自连而成;b3.2':b1.14'经HindI和XhoI部分酶解所得片段插入经同样的两个酶酶解的pBLCAT3而成。b3.4':b1.15'经Hind和XhoI部分酶解所得片段插入经同样的两种酶酶解的pBLCAT3而成 。
, http://www.100md.com
1.2.2 质粒转染:2000r/min离心5min收集2×107个Jurkat细胞,用10mL 37 ℃预热的STBS溶液[(含mmol/L):Tris-HCl 125,NaCl 137,KCl 5,NaH2PO4 0.6,CaCl2 0.7,MgCl2 0.5]洗一次,将10mg质粒DNA,1mL STBS溶液及0.05mL DEAE-Dextron(10g/L STBS)的混合液重悬细胞,37 ℃温育1h(期间每隔15min轻轻混匀一次)后,滴加110mL DMSO,迅速混匀,2min30s后加15mL STBS稀释(相当于终止反应),1500r/min离心5min,细胞沉淀依次用10mL STBS、10mL无血清RPMI 1640培养基各洗一次,加完全培养基重悬细胞,分至6孔或12孔培养板,37 ℃、5% CO2培养48h后裂解细胞。细胞如果要用PHA刺激,则在收细胞前18h加入PHA(无血清1640配制)至终浓度7.5mg/L。若要进行热刺激,则在收细胞前1h将细胞在42 ℃水浴中热休克1h。
, http://www.100md.com
1.2.3 报告基因活性的检测
1.2.3.1 氯霉素乙酰基转移酶(CAT)活性的测定:收集转染的细胞,用冰预冷的PBS洗两遍,加100mL 250mmol/L的Tris-HCl(pH 7.8)重悬细胞,在液氮和37 ℃水浴中反复冻融三次,在4 ℃以12 000r/min离心10min,取30mL上轻液,加入20mL 100mmol/L的Tris-HCl(pH 7.8),65 ℃灭活脱乙酰基酶15min。然后加入200mL CAT反应液(125mmol/L Tris-HCl, pH 7.8,1.25mmol/L氯霉素, 0.1mCi 3H标记的乙酰辅酶A),转移至液闪瓶中,上面覆盖5mL闪烁液,37 ℃扩散16h后测cpm值。然后将按BCA法(PIERCE公司)所测得的蛋白浓度,校正相应裂解液的cpm值,称相对CAT活性。
1.2.3.2 荧光素酶(Luc)活性的测定收集转染的细胞,用预冷的PBS洗两遍,加入100mL 1×细胞裂解液(lysis buffer),充分混匀,室温裂解 15~20min后,4 ℃,12000r/min离心10min,取上轻20mL至发光管中,在荧光检测仪(Monolight 2010)中进行检测,所得的值用 b3.1中同样的测定方法进行校正后即为相对 Luc活性。
, 百拇医药
2 结果
2.1 CRE和AP-1元件删切报告基因质粒的构建
为了探讨CRE利 AP-l元件在hsp90β基因表达调控中的作用,我们构建了对这两个元件进行不同删切的含有hsp90β基因调控序列的报告基因质粒(见图1)。为研究方便,我们将调控序列连接到两种报告基问CAT和Luc上。其中β3.1和β1.11是含有全长调控序列的报告基因质粒 ;β3.2和β1.14同 时含有CRE和AP-1元件;β3.4和b1.15只含有AP-1元件;β3.2'和b1.14'只含有CRE元件;β3.4'和β1.15'中两种元件均被删除。这些质粒均经限制性内切酶酶谱鉴定。
2.2 CRE和 AP-l元件参与 hsp90β基因的高组成性表达
将上述系列删切质粒分别转染Jurkat细胞,通过分析各组细胞裂解液中报告基因Luc的活性,我们发现:单独删除CRE(βl.15)或单独删除AP-1(β1.14')元件,hsp90β基因的组成性表达均稍稍有所降低,但变化不明显;但当同时删除这两个元件时(β1.15'),组成性表达降低了40%(图2)。这表明,CRE和AP-1元件在hsp90β基因的高组成性表达中起着重要作用。
, 百拇医药
2.3 CRE和 AP-l元件介导 PHA对 hsp90β基因的激活表达
通过对Luc活性的分析,我们发现:保留CRE和 AP-l元件中的任何一个,hsp90β基因的表达均可被PHA所激活,只是激活倍数比同时含有这两个元件时有所降低;但当同时删除这两个元件时,PHA便不能激活hsp90β基因的表达(图2)。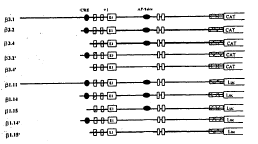
图1 CRE和AP-1邞件序列删切质粒的构建
Fig1 Plasmids construction of deleting CRE and AP-1 elements in the regulatory sequences of hsp90β gene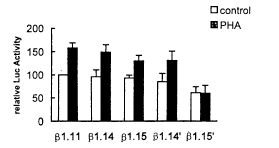
, 百拇医药
图2 CRE和AP-1元件对hsp90β基因组成性和
PHA激活表达的影响
Fig2 Effects of CRE and AP-1 elements in the constitutive and PHA activated expression of hsp90β gene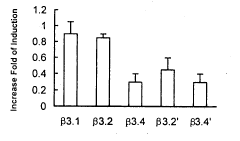
图3 CRE和AP-1元件对hsp90β基因热诱导表达的影响
Fig3 Effects of CRE and AP-1 elements in the heat
inducible expression of hsp90β gene
, 百拇医药
2.4 CRE和AP-1元件参与 hsp90β基因的热诱导表达
由于Luciferase对热不稳定,我们将含有CAT报告基因的质粒转染Jurkat细胞。通过分析CAT的活性,我们发现:含有hsp90β基因全长调控序列(-1039~+1531)质粒β3.1的热诱导倍数能增加90%;单独删除 CRE元件(β3.4)、单独删除AP-1元件(β3.2')或同时删除这两个元件(β3.4')时,热诱导倍数分别降低了67%、50%和67%(图3)。这说明CRE和AP-1都不同程度地参与了hsp90β基因的热诱导表达。
3 讨论
结合上述结果,我们认为:(1)CRE和AP-1元件参与了hsp90b基因的高组成性表达。(2) CRE和 AP-1是 PHA激活hsp90β基因表达的反应元件,它们可以共同或单独介导PHA对hsp90β基因的激活表达。(3)CRE和 AP-1均参与了hsp90β基因的热诱导表达,在热诱导中CRE的作用要比AP-1的大。
, http://www.100md.com
在上述的结果中,我们发现,在hsp90β基因的组成性表达中,CRE和AP-1元件对hsp90β基因的高组成性表达所起的作用有些特殊,即只要存在其中任何一个元件,组成性表达均无明显变化。对于这一现象,我们推测CRE可能介导从+1位点起始的转录,而AP-1可以介导从第一内含子3'端起始的转录。本组已经证明了在第一内含子3'端有多个转录起始位点[4]。hsp90β基因PHA的激活表达情况与组成性表达相似,我们推测:(1)PHA激活通路可能有两条,这两条通路的最终反应元件分别是CRE和 AP-1元件。(2)由于这两个元件的结合蛋白CREB和JUN/FOS已被证明可以有选择地组成异源二聚体[5]。因此,这些异源二聚体是不是有可能和该元件中的任何一个均有结合活性,最终促进基因的转录?另外还有别的实验也证明了CRE和 AP-1共同介导一些基因的表达[8,9]。在hsp90β基因的热诱导表达中,这两个元件所起的作用使我们推测 CREB和 JUN/FOS与热休克因子 HSF、 P300/CBP等之间很可能存在一些重要的联系。目前有关的研究正在进行之中。
, http://www.100md.com
基金项目:自然科学基金39770169,39930050,和863课题102-11-02-05资助
[1] 沈珝琲,吴宁华,程小款,等.热休克与ConA激活对T淋巴细胞的双重作用[J].生物化学杂志,1986,2(5):35-39.
[2] Hicky E,Brandon SE,Smale G,et al.Sequence and regulation of a gene encoding a human 89-kilodalton heat shock protein[J].Mol Cell Biol,1989,9:2615-2626.
[3] Rebbe NF,Hickman WS,Ley TJ,et al.Nucleotide sequence of a cDNA for a member of the human 90-kDa heat-shock protein family[J].Gene,1987,53:235-245.
, 百拇医药
[4] Shen YF,Liu JH,Wang XZ,et al.Essential roles of the first intron in transcription regulation of hsp90b gene[J].FEBS letters,1997,413:92-98.
[5] Hai T,Curran T.Cross-family dimerizaion of transcription factors Fos: Jun and ATF/CREB alters DNA Binding specificity[J].PNAS USA,1991,88:3720-3724.
[6] Parker D,Ferreri K,Nakajima T,et al.Phosphorylation of CREB at Ser-133 induces complex formation with CREB-Binding Protein via a direct mechanism[J].Mol Cell Biol,1996,16(2):694:703.
, 百拇医药
[7] Shikama N,Lyon J,La Thangue NB.The P300/CBP family: intergrating signals with transcription factors and chomatin[J].Trends in Cell Biology,1997,7:230-236.
[8] Ishiguro N,Brown GD,Ishizu A,et al.The regulation of murine H-2Dd expression by activation transcription factor 1 and cAMP response element binding protein[J].J Immunol,1998,160(12):5907-5914.
[9] Cirillo G,Casalino L,Vallone D,et al.Role of distinc mitogen-activated protein kinase pathways and cooperation between Ets-2,ATF-2,and Jun family menbers in human urokinase-type plasminogen activater gene induction by interleukin-1 and tetradecanoyl phorbol acetate[J].Mol Cell Biol,1999,19(9):6240-6252.
收稿日期:2000-01-01; 修订日期:2000-02-25, 百拇医药
单位:刘秉胜(中国医学科学院基础医学研究所 中国协和医科大学基础医学院 医学分子生物学国家重点实验室,北京100005);吴宁华(中国医学科学院基础医学研究所 中国协和医科大学基础医学院 医学分子生物学国家重点实验室,北京100005);沈珝琲(中国医学科学院基础医学研究所 中国协和医科大学基础医学院 医学分子生物学国家重点实验室,北京100005)
关键词:CRE元件;AP-1元件;hsp90b基因;基因表达
基础医学与临床000306
摘要:为了进一步阐明人热休克蛋白90b(hsp90b)基因的表达调控机制,本文对hsp90b基因调控序列中的CRE和 AP-1元件进行了研究。构建了含有这两个元件的系列删切报告基因质粒,通过转染 Jurkat细胞,检测并分析报告基因的活性。结果表明,CRE和 AP-1元件均不同程度地参与了hsp90b基因的组成性、PHA激活和热诱导表达。
, http://www.100md.com
中图分类号:Q786 文献标识码:A 文章编号:1001-6352(2000)03-0028-04
CRE and AP-1 elements in the expression of human hsp90b gene
LIU Bing-sheng,WU Ning-hua,SHEN Yu-fei
(National Laboratory of Medical Molecular Biology,Institute of Basic Medical Sciences,Chinese Academy
of Medical Science&Peking Union Medical College,Beijing100005,China)
, 百拇医药 Abstract:Several reporter plasmids with deleting CRE and AP-1 elements in the regulation sequences of hsp90b gene were constructed and transfected into Jurkat cells.Then the reporter genes' activity was detected.The results show that CRE and AP-1 elements were critical to the high constitutive expression of hsp90b gene; in the PHA activated expression,CRE or AP-1 elements play synergistic role; CRE and AP-1 elements also partially mediate in the heat shock inducible expression of hsp90b gene.
, http://www.100md.com
Key words: CRE; AP-1 element; hsp90b gene; gene expression
hsp90基因是热休克蛋白基因家族的成员。与其它热休克蛋白基因(如hsp70)不同hsp90基因通常在细胞内就有较高水平的组成性表达,并可进一步被热和丝裂原诱导[1]。 hsp90基因有a和b两个拷贝[2,3]。本组对hsp90b基因的表达顺式调节机理进行了研究。研究表明,上游启动子元件、核心启动子和第一外显子或第一内含子对于高组成性表达起重要作用;在热诱导表达中,第一内含子中两个典型的 HSE起了非常重要的作用[4]。我们在分析这些区域的顺式元件分布时,发现在上游启动区-127/-120bp处为环腺苷酸反应元件 (cAMP responsive element,简称 CRE)或称为转录激活因子结合位点 (the activating transcription factor,简称 ATF),同时在第一内含子+484/+489bp处有一 AP-1(the activator protein l)元件(或TRE元件, the phorbol 12-0-tetradecanoate 13-acetate responsive element,简称TRE)。 CRE元件的序列是:TGACGTCA。AP-1元件的序列是:TGACTCA。这两种元件可分别被CRE结合蛋白 CREB家族成员和 AP-1识别蛋白Jun/Fos所识别。研究表明这些转录因子均与信号传导有关[5~7]。因此,为了进一步深入了解 hsp90b基因的表达调控机理,我们对这两个元件在hsp90b基因表达调控中的作用进行了研究。
, 百拇医药
1 材料与方法
1.1 材料
1.1.1 主要试剂:Hinc、XhoI、BamHI、SmaI等限制性内切酶和T4 DNA连接酶为Boehinger Mannheim公司产品;其它限制性内切酶及DNA分子量标准均为协和友谊公司产品;PHA为Sigma公司产品;细胞裂解及荧光素酶检测试剂盒为Promega公司产品;3H-乙酰辅酶A为Amersham公司产品;BCA 蛋白分析试剂为Pierce公司产品。
1.1.2 质粒与细胞株:含有hsp90b基因调控序列-1039至+1531bp的CAT基因报告质粒b3.1和荧光素酶基因报告质粒b1.11由本课题组提供;Jurkat细胞为本课题组保存的细胞株。
1.2 方法
1.2.1 CRE和AP-1元件系缺失质粒的构建:b1.14经BamHI和XhoI部分酶解所得片段-173/+1351插入PMC/Luc报告质粒而建成;b1.15:由b1.11经Hinc和XhoI部分酶切所得片段-91/+1531插入经SmaI和XhoI酶切的PMC/Luc报告质粒建成;b1.14':由b1.14经SmaI和MluI酶解所得大片段用Klenow酶补平后自连而成;b1.15'由b1.15经SmaI和MluI酶解所得大片段用Klenow补平后自连而成。b3.2:b3.1经BamHI和XhoI部分酶解所得片段-173/+1531插入被SmaI和XhoI酶解的pBLCAT3报告质粒而建成;b3.4:b3.1经PstI完全酶解,Klenow酶补平后经Hinc部分酶解所得大片段自连而成;b3.2':b1.14'经HindI和XhoI部分酶解所得片段插入经同样的两个酶酶解的pBLCAT3而成。b3.4':b1.15'经Hind和XhoI部分酶解所得片段插入经同样的两种酶酶解的pBLCAT3而成 。
, http://www.100md.com
1.2.2 质粒转染:2000r/min离心5min收集2×107个Jurkat细胞,用10mL 37 ℃预热的STBS溶液[(含mmol/L):Tris-HCl 125,NaCl 137,KCl 5,NaH2PO4 0.6,CaCl2 0.7,MgCl2 0.5]洗一次,将10mg质粒DNA,1mL STBS溶液及0.05mL DEAE-Dextron(10g/L STBS)的混合液重悬细胞,37 ℃温育1h(期间每隔15min轻轻混匀一次)后,滴加110mL DMSO,迅速混匀,2min30s后加15mL STBS稀释(相当于终止反应),1500r/min离心5min,细胞沉淀依次用10mL STBS、10mL无血清RPMI 1640培养基各洗一次,加完全培养基重悬细胞,分至6孔或12孔培养板,37 ℃、5% CO2培养48h后裂解细胞。细胞如果要用PHA刺激,则在收细胞前18h加入PHA(无血清1640配制)至终浓度7.5mg/L。若要进行热刺激,则在收细胞前1h将细胞在42 ℃水浴中热休克1h。
, http://www.100md.com
1.2.3 报告基因活性的检测
1.2.3.1 氯霉素乙酰基转移酶(CAT)活性的测定:收集转染的细胞,用冰预冷的PBS洗两遍,加100mL 250mmol/L的Tris-HCl(pH 7.8)重悬细胞,在液氮和37 ℃水浴中反复冻融三次,在4 ℃以12 000r/min离心10min,取30mL上轻液,加入20mL 100mmol/L的Tris-HCl(pH 7.8),65 ℃灭活脱乙酰基酶15min。然后加入200mL CAT反应液(125mmol/L Tris-HCl, pH 7.8,1.25mmol/L氯霉素, 0.1mCi 3H标记的乙酰辅酶A),转移至液闪瓶中,上面覆盖5mL闪烁液,37 ℃扩散16h后测cpm值。然后将按BCA法(PIERCE公司)所测得的蛋白浓度,校正相应裂解液的cpm值,称相对CAT活性。
1.2.3.2 荧光素酶(Luc)活性的测定收集转染的细胞,用预冷的PBS洗两遍,加入100mL 1×细胞裂解液(lysis buffer),充分混匀,室温裂解 15~20min后,4 ℃,12000r/min离心10min,取上轻20mL至发光管中,在荧光检测仪(Monolight 2010)中进行检测,所得的值用 b3.1中同样的测定方法进行校正后即为相对 Luc活性。
, 百拇医药
2 结果
2.1 CRE和AP-1元件删切报告基因质粒的构建
为了探讨CRE利 AP-l元件在hsp90β基因表达调控中的作用,我们构建了对这两个元件进行不同删切的含有hsp90β基因调控序列的报告基因质粒(见图1)。为研究方便,我们将调控序列连接到两种报告基问CAT和Luc上。其中β3.1和β1.11是含有全长调控序列的报告基因质粒 ;β3.2和β1.14同 时含有CRE和AP-1元件;β3.4和b1.15只含有AP-1元件;β3.2'和b1.14'只含有CRE元件;β3.4'和β1.15'中两种元件均被删除。这些质粒均经限制性内切酶酶谱鉴定。
2.2 CRE和 AP-l元件参与 hsp90β基因的高组成性表达
将上述系列删切质粒分别转染Jurkat细胞,通过分析各组细胞裂解液中报告基因Luc的活性,我们发现:单独删除CRE(βl.15)或单独删除AP-1(β1.14')元件,hsp90β基因的组成性表达均稍稍有所降低,但变化不明显;但当同时删除这两个元件时(β1.15'),组成性表达降低了40%(图2)。这表明,CRE和AP-1元件在hsp90β基因的高组成性表达中起着重要作用。
, 百拇医药
2.3 CRE和 AP-l元件介导 PHA对 hsp90β基因的激活表达
通过对Luc活性的分析,我们发现:保留CRE和 AP-l元件中的任何一个,hsp90β基因的表达均可被PHA所激活,只是激活倍数比同时含有这两个元件时有所降低;但当同时删除这两个元件时,PHA便不能激活hsp90β基因的表达(图2)。
图1 CRE和AP-1邞件序列删切质粒的构建
Fig1 Plasmids construction of deleting CRE and AP-1 elements in the regulatory sequences of hsp90β gene
, 百拇医药
图2 CRE和AP-1元件对hsp90β基因组成性和
PHA激活表达的影响
Fig2 Effects of CRE and AP-1 elements in the constitutive and PHA activated expression of hsp90β gene
图3 CRE和AP-1元件对hsp90β基因热诱导表达的影响
Fig3 Effects of CRE and AP-1 elements in the heat
inducible expression of hsp90β gene
, 百拇医药
2.4 CRE和AP-1元件参与 hsp90β基因的热诱导表达
由于Luciferase对热不稳定,我们将含有CAT报告基因的质粒转染Jurkat细胞。通过分析CAT的活性,我们发现:含有hsp90β基因全长调控序列(-1039~+1531)质粒β3.1的热诱导倍数能增加90%;单独删除 CRE元件(β3.4)、单独删除AP-1元件(β3.2')或同时删除这两个元件(β3.4')时,热诱导倍数分别降低了67%、50%和67%(图3)。这说明CRE和AP-1都不同程度地参与了hsp90β基因的热诱导表达。
3 讨论
结合上述结果,我们认为:(1)CRE和AP-1元件参与了hsp90b基因的高组成性表达。(2) CRE和 AP-1是 PHA激活hsp90β基因表达的反应元件,它们可以共同或单独介导PHA对hsp90β基因的激活表达。(3)CRE和 AP-1均参与了hsp90β基因的热诱导表达,在热诱导中CRE的作用要比AP-1的大。
, http://www.100md.com
在上述的结果中,我们发现,在hsp90β基因的组成性表达中,CRE和AP-1元件对hsp90β基因的高组成性表达所起的作用有些特殊,即只要存在其中任何一个元件,组成性表达均无明显变化。对于这一现象,我们推测CRE可能介导从+1位点起始的转录,而AP-1可以介导从第一内含子3'端起始的转录。本组已经证明了在第一内含子3'端有多个转录起始位点[4]。hsp90β基因PHA的激活表达情况与组成性表达相似,我们推测:(1)PHA激活通路可能有两条,这两条通路的最终反应元件分别是CRE和 AP-1元件。(2)由于这两个元件的结合蛋白CREB和JUN/FOS已被证明可以有选择地组成异源二聚体[5]。因此,这些异源二聚体是不是有可能和该元件中的任何一个均有结合活性,最终促进基因的转录?另外还有别的实验也证明了CRE和 AP-1共同介导一些基因的表达[8,9]。在hsp90β基因的热诱导表达中,这两个元件所起的作用使我们推测 CREB和 JUN/FOS与热休克因子 HSF、 P300/CBP等之间很可能存在一些重要的联系。目前有关的研究正在进行之中。
, http://www.100md.com
基金项目:自然科学基金39770169,39930050,和863课题102-11-02-05资助
[1] 沈珝琲,吴宁华,程小款,等.热休克与ConA激活对T淋巴细胞的双重作用[J].生物化学杂志,1986,2(5):35-39.
[2] Hicky E,Brandon SE,Smale G,et al.Sequence and regulation of a gene encoding a human 89-kilodalton heat shock protein[J].Mol Cell Biol,1989,9:2615-2626.
[3] Rebbe NF,Hickman WS,Ley TJ,et al.Nucleotide sequence of a cDNA for a member of the human 90-kDa heat-shock protein family[J].Gene,1987,53:235-245.
, 百拇医药
[4] Shen YF,Liu JH,Wang XZ,et al.Essential roles of the first intron in transcription regulation of hsp90b gene[J].FEBS letters,1997,413:92-98.
[5] Hai T,Curran T.Cross-family dimerizaion of transcription factors Fos: Jun and ATF/CREB alters DNA Binding specificity[J].PNAS USA,1991,88:3720-3724.
[6] Parker D,Ferreri K,Nakajima T,et al.Phosphorylation of CREB at Ser-133 induces complex formation with CREB-Binding Protein via a direct mechanism[J].Mol Cell Biol,1996,16(2):694:703.
, 百拇医药
[7] Shikama N,Lyon J,La Thangue NB.The P300/CBP family: intergrating signals with transcription factors and chomatin[J].Trends in Cell Biology,1997,7:230-236.
[8] Ishiguro N,Brown GD,Ishizu A,et al.The regulation of murine H-2Dd expression by activation transcription factor 1 and cAMP response element binding protein[J].J Immunol,1998,160(12):5907-5914.
[9] Cirillo G,Casalino L,Vallone D,et al.Role of distinc mitogen-activated protein kinase pathways and cooperation between Ets-2,ATF-2,and Jun family menbers in human urokinase-type plasminogen activater gene induction by interleukin-1 and tetradecanoyl phorbol acetate[J].Mol Cell Biol,1999,19(9):6240-6252.
收稿日期:2000-01-01; 修订日期:2000-02-25, 百拇医药