肿瘤抑制基因p53的生物学功能研究进展和意义
作者:朱明华
单位:朱明华(上海第二军医大学病理学教研室 200433)
关键词:
中华病理学杂志000122 肿瘤抑制基因p53为细胞癌基因中研究最为广泛和深入的基因之一,有关的研究论文平均每年超过1 000篇,并为Science杂志评为1993年度的明星分子[1]。经过多年的研究,对p53基因的结构已有了明确的认识,对p53基因的功能研究也取得重要进展。p53与多种基因之间均存在相互调节的作用,但对其中大部分的调节机制尚不明确。目前细胞癌基因的研究已由单一基因发展到对多基因协同作用和相互调节机制的研究阶段。p53基因的异常几乎存在于人类的所有肿瘤,故对其作用机制的研究显得尤为重要。但对于p53基因及其表达产物在正常细胞周期中的作用机制、基因间的调节及在肿瘤发生中的具体作用,诸如是通过何种确切途径去启动生长停滞和细胞凋亡?细胞DNA损伤时调动p53积聚的信号有那些等方面还存在许多有待解决的问题。
, http://www.100md.com
一、p53与p21WAF1/CIP1
p21WAF1/CIP1基因编码产物是一种细胞周期蛋白依赖性激酶抑制蛋白(cyclin-dependent kinase inhibitor,CDKI)。它通过调控细胞周期的进程,参与细胞的生长、分化、衰老及死亡。在p21编码区上游2.4 kb和大于8 kb处有2个p53结合区,在上游75 bp处有一个弱p53结合区[2],实验表明,在p53+/+的细胞中,DNA损伤时常有p53和p21的高表达,而在p53-/-细胞,则无p21表达,细胞也无G1期的停滞。p21作为p53的下游激活产物,执行p53的部分功能。当细胞DNA损伤后,p53蛋白积聚,使p21基因表达上调,细胞阻滞于G1期,以赢得时间,在细胞进入S期前修复损伤的DNA。p53和p21的相互调节对于使损伤的细胞阻滞于G1期是必需的。最近Vogelstein等[3]又报道了新的研究结果,认为细胞在DNA损伤后,p53和p21的调节作用对于细胞阻滞于G2期也是必需的。p53和p21参与细胞DNA损伤修复机制可参阅图1。
, 百拇医药
二、p53与mdm2及ARF
mdm2为一种进化保守基因,具有转录因子功能,野生型(wt)或突变型(mt)的p53均可与mdm2基因产物结合,使p53介导的反式激活、抑制细胞增殖和诱导凋亡的作用受抑制,解除细胞G1期的阻滞并重新进入细胞周期。mdm2蛋白为p53基因激活的靶点,目前认为野生型p53基因对控制细胞周期,维持细胞遗传稳定性具有重要作用,并认为可能是通过GADD45(growth arrest and DNA damage inducible gene)和 mdm2行使调节作用。p53蛋白与mdm2蛋白结合构成寡二聚体复合物,mdm2过度表达则抑制p53的转录活性。抑制作用是通过mdm2与p53蛋白酸性活化区直接结合实现的。无论细胞内mdm2是否扩增,野生型p53的表达都能强烈影响mdm2蛋白在相同细胞中的表达。二者不但在调节细胞分裂、增殖方面存在一种相互调节的机制,而且非常复杂。
Kamijo等[4]和Velculescu等[5]报道,肿瘤抑制基因INK4a-ARF的编码蛋白p19ARF与p53、mdm2均存在功能上的相互调节作用,ARF与p53之间存在直接的相互作用关系,通过提高p53功能稳定性或其他尚未知的机制调节细胞功能。
, http://www.100md.com
图1 p53和p21参与细胞损伤修复的分子途径(参考文献[5])
三、p53与最近发现的几种基因产物的关系
Schmale等[6]于1997年从大鼠的表皮细胞中克隆了一种基因,命名为Ket,在C-末端的第498~606氨基酸残基部位与人p53高度同源。该基因具有p53所具备的生物学功能,属于一种分化、发育相关的基因,可能还具有组织特异性分化的功能。Ket基因的出现,为p53基因的研究又提出了新的课题。
Nishimori等[7]克隆了一种含1 584个氨基酸残基的基因,命名为BAI1,特异性在脑组织中表达,可以抑制胶质瘤组织中的血管生成。目前发现,BAI1是p53调节的靶基因。
Israeli等[8]克隆了一种长7.5 kb的基因PAG608。该基因编码一种核内的锌指蛋白,中等表达量即可使细胞核形态改变,过度表达时能诱导细胞凋亡。研究发现,PAG608在细胞DNA损伤时mRNA表达和蛋白质表达增加都依赖于p53的调控,为一种p53的靶基因。
, 百拇医药
Simons等[9]于1997年报道了命名为PACT的基因克隆和特征。该基因的克隆是以p53作探针从表达文库中得到,编码相对分子质量250 000的核内蛋白。应用免疫沉淀和免疫吸附试验,证明PACT蛋白能与野生型p53和Rb蛋白结合,与p53蛋白结合后,能影响p53基因的特异性DNA结合功能。有关PACT基因及其产物的生物学功能仍有待更多的研究探索。
Osada等[10]在1998年克隆到一个新基因命名为p51。其后又发现几个与p53高度同源的新基因如p73A、p73B、p51A、p51B、p63等。这些基因与p53除了结构上高度同源外,在功能上也存在非常复杂的联系,所以把这一类基因称作为p53基因家族,其生物学功能和意义以及在肿瘤发生中的作用正在研究之中。
四、p53与DNA肿瘤病毒蛋白的相互作用
p53基因产物为核内磷酸化蛋白,具有蛋白质-蛋白质和DNA-蛋白质结合的功能。p53蛋白不仅可与双链DNA结合,也具有与单链DNA结合的能力,除了能与细胞内某些基因产物蛋白结合,还可与一些肿瘤病毒转化蛋白结合并使p53功能失活。 研究认为除p53突变外,肿瘤病毒蛋白与p53结合是p53改变与肿瘤发生关系中最主要的原因。目前已报道的能与p53结合的细胞内蛋白有CBF、E6-AP、ERCC3、HSP70、mdm2、RPA、SP1、TAFⅡ40、TAFⅡ60、TBP和WT1等;能与p53结合的DNA肿瘤病毒蛋白有腺病毒E1B、EBNA5、HPV-E6、SV40T和HBxAg。p53基因与肿瘤病毒相互作用的生物学机制如图2所示。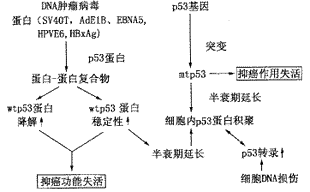
, http://www.100md.com
注:方框内为最终结果
图2 p53与肿瘤病毒蛋白相互作用与
肿瘤发生机制(参考文献[5])
p53蛋白与SV40 T、腺病毒E1B、EBNA5和HPV6的相互关系及在肿瘤发生中的作用,已有许多报道,现介绍乙肝病毒X蛋白与p53的关系及其在肝癌发生中的作用机制。
乙型肝炎病毒(HBV)为一种小型的DNA病毒,HBV慢性感染者发生肝癌的危险性比无感染者高200倍左右。虽经多年研究,HBV致肝癌的确切机制仍不明确。HBV X基因为4个开放阅读框架中最小的1个,其编码产物HBxAg具有反式激活作用,与肝癌的发生具有密切关系。肝癌中p53突变频率是肿瘤基因突变较高的一种。在一些与HBV感染相关的肝癌中,发现存在p53蛋白细胞内积聚,但p53基因突变率则较低,提示HBxAg可能具有与其他DNA肿瘤病毒编码产物类似的作用,即与p53之间存在蛋白-蛋白结合的关系。为证实这一假设,我们[11,12]应用免疫沉淀和细胞内掺入的方法,于1993年报道了HBxAg与p53存在蛋白-蛋白结合。继之其他实验室进一步研究证实,HBV是继SV40、腺病毒、EBV和HPV之后,第5个病毒编码产物能与p53蛋白结合的DNA病毒。这一现象不仅存在人HBV,也存在于WHV[13]。进一步对HBV基因组进行分析发现,在HBV增强子Ⅰ(enhancer Ⅰ)上游的1047~1059 bp区域,存在经典的p53基因DNA-蛋白质结合位点类似的基因序列,即2个TGCCT正向重复序列。经凝胶电泳迁移率等实验证实,这一序列可特异性地与p53蛋白结合[14]。以上研究初步表明,在HBV感染相关性肝癌中,HBV与p53之间不仅存在蛋白-蛋白(HBxAg-p53)的结合,还存在DNA-蛋白质(HBV-p53)的结合,导致p53蛋白在细胞内积聚,正常的负调节功能丧失。p53在HBV相关性肝癌发生中的可能作用机制如图3所示。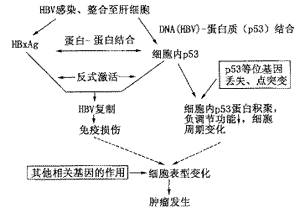
, http://www.100md.com
注:方框内指病毒以外的相关因素
图3 p53与HBV相关性肝癌发生的可能机制
综上所述,关于抑癌基因p53与肿瘤关系,近年来较注重于p53功能、基因间调节、与其他因子相互关系等方面的研究。随着研究的不断深入,进一步加深对p53基因生物学功能和在肿瘤发生发展过程中作用的理解,将更有助于肿瘤的诊断、治疗和预后评估。
参考文献:
[1]Koshland DE Jr. Molecular of the year .Science,1993,262:1953.
[2]El-Deiry WS, Tokino T, Velculescu VE, et al. WAF1, a potential mediator of p53 tumor suppression. Cell ,1993,75:817-825.
, 百拇医药
[3]Bunz F, Dutriaux A, Lengauer C, et al. Requirement for p53 and p21 to sustain G2 arrest after DNA damage. Science,1998,282:1497-1501.
[4]Kamijo T, Weber JD ,Zambetti G, et al. Functioal and physical interactions of the ARF tumor suppressor with p53 and mdm2. Proc Natl Acad Sci USA,1998,95:8292-8297.
[5]Velculescu VE, EL-Deiry WS. Biological and clinical importance of the p53 tumor suppressor gene. Clin Chem, 1996,42:858-869.
, 百拇医药
[6]Schmale H, Bamberger C. A novel protein with strong homology to the tumor suppressor p53. Oncogene ,1997,15:1363-1367.
[7]Nishimori H, Shiratsuchi T, Urano T, et al. A novel brain-specific p53-target gene,BAI1,containing thrombospondin type 1 repeats inhibits experimental angiogenosis. Oncogene,1997,15:2145-2150.
[8]Israeli D, Tessler E, Haupt Y, et al. A novel p53-inducible gene,PAG608, encodes a nuclear zinc finger protein whose overexpression promotes apoptosis. EMBO J,1997,16:4384-4392.
, 百拇医药
[9]Simons A, Melamed-Bessudo C,Wolkowicz R,et al. PACT:cloning and characterization of cellular p53 binding protein that interacts with Rb. Oncogene,1997,14:145-155.
[10]Kaelin WJ. The emerging p53 family. J Natl Cancer Inst,1999,91:594- 598.
[11]朱明华,Feitelgon MA, London WT. HBxAg和p53蛋白结合在原发性肝癌发 生中的意义.中华医学杂志,1993,73:325-327.
[12]Feitelson MA, Zhu MH, Duan LX ,et al. Hepatitis B X antigen and p53 are associated in vitro and in liver tissues from patients with primary hepatocellular Carcinoma. Oncogene, 1993,8:1109-1117.
, 百拇医药
[13]Feitelson MA,Ranganathan PN,Clayton MM, et al. Partial characterization of woodchuck tumor suppressor ,p53, and its interaction with woodchuck hepatitis virus X antigen in hepatocarcinogenesis. Oncogene, 1997,15:327-336.
[14]朱明华,Duan LX,Feitelson MA, et al. 乙型肝炎病毒与肿瘤抑制基因p53 相互作用的生物学功能及意义.病毒学报,1998,14:10-15.
收稿日期:1999-06-21, 百拇医药
单位:朱明华(上海第二军医大学病理学教研室 200433)
关键词:
中华病理学杂志000122 肿瘤抑制基因p53为细胞癌基因中研究最为广泛和深入的基因之一,有关的研究论文平均每年超过1 000篇,并为Science杂志评为1993年度的明星分子[1]。经过多年的研究,对p53基因的结构已有了明确的认识,对p53基因的功能研究也取得重要进展。p53与多种基因之间均存在相互调节的作用,但对其中大部分的调节机制尚不明确。目前细胞癌基因的研究已由单一基因发展到对多基因协同作用和相互调节机制的研究阶段。p53基因的异常几乎存在于人类的所有肿瘤,故对其作用机制的研究显得尤为重要。但对于p53基因及其表达产物在正常细胞周期中的作用机制、基因间的调节及在肿瘤发生中的具体作用,诸如是通过何种确切途径去启动生长停滞和细胞凋亡?细胞DNA损伤时调动p53积聚的信号有那些等方面还存在许多有待解决的问题。
, http://www.100md.com
一、p53与p21WAF1/CIP1
p21WAF1/CIP1基因编码产物是一种细胞周期蛋白依赖性激酶抑制蛋白(cyclin-dependent kinase inhibitor,CDKI)。它通过调控细胞周期的进程,参与细胞的生长、分化、衰老及死亡。在p21编码区上游2.4 kb和大于8 kb处有2个p53结合区,在上游75 bp处有一个弱p53结合区[2],实验表明,在p53+/+的细胞中,DNA损伤时常有p53和p21的高表达,而在p53-/-细胞,则无p21表达,细胞也无G1期的停滞。p21作为p53的下游激活产物,执行p53的部分功能。当细胞DNA损伤后,p53蛋白积聚,使p21基因表达上调,细胞阻滞于G1期,以赢得时间,在细胞进入S期前修复损伤的DNA。p53和p21的相互调节对于使损伤的细胞阻滞于G1期是必需的。最近Vogelstein等[3]又报道了新的研究结果,认为细胞在DNA损伤后,p53和p21的调节作用对于细胞阻滞于G2期也是必需的。p53和p21参与细胞DNA损伤修复机制可参阅图1。
, 百拇医药
二、p53与mdm2及ARF
mdm2为一种进化保守基因,具有转录因子功能,野生型(wt)或突变型(mt)的p53均可与mdm2基因产物结合,使p53介导的反式激活、抑制细胞增殖和诱导凋亡的作用受抑制,解除细胞G1期的阻滞并重新进入细胞周期。mdm2蛋白为p53基因激活的靶点,目前认为野生型p53基因对控制细胞周期,维持细胞遗传稳定性具有重要作用,并认为可能是通过GADD45(growth arrest and DNA damage inducible gene)和 mdm2行使调节作用。p53蛋白与mdm2蛋白结合构成寡二聚体复合物,mdm2过度表达则抑制p53的转录活性。抑制作用是通过mdm2与p53蛋白酸性活化区直接结合实现的。无论细胞内mdm2是否扩增,野生型p53的表达都能强烈影响mdm2蛋白在相同细胞中的表达。二者不但在调节细胞分裂、增殖方面存在一种相互调节的机制,而且非常复杂。
Kamijo等[4]和Velculescu等[5]报道,肿瘤抑制基因INK4a-ARF的编码蛋白p19ARF与p53、mdm2均存在功能上的相互调节作用,ARF与p53之间存在直接的相互作用关系,通过提高p53功能稳定性或其他尚未知的机制调节细胞功能。
, http://www.100md.com
图1 p53和p21参与细胞损伤修复的分子途径(参考文献[5])
三、p53与最近发现的几种基因产物的关系
Schmale等[6]于1997年从大鼠的表皮细胞中克隆了一种基因,命名为Ket,在C-末端的第498~606氨基酸残基部位与人p53高度同源。该基因具有p53所具备的生物学功能,属于一种分化、发育相关的基因,可能还具有组织特异性分化的功能。Ket基因的出现,为p53基因的研究又提出了新的课题。
Nishimori等[7]克隆了一种含1 584个氨基酸残基的基因,命名为BAI1,特异性在脑组织中表达,可以抑制胶质瘤组织中的血管生成。目前发现,BAI1是p53调节的靶基因。
Israeli等[8]克隆了一种长7.5 kb的基因PAG608。该基因编码一种核内的锌指蛋白,中等表达量即可使细胞核形态改变,过度表达时能诱导细胞凋亡。研究发现,PAG608在细胞DNA损伤时mRNA表达和蛋白质表达增加都依赖于p53的调控,为一种p53的靶基因。
, 百拇医药
Simons等[9]于1997年报道了命名为PACT的基因克隆和特征。该基因的克隆是以p53作探针从表达文库中得到,编码相对分子质量250 000的核内蛋白。应用免疫沉淀和免疫吸附试验,证明PACT蛋白能与野生型p53和Rb蛋白结合,与p53蛋白结合后,能影响p53基因的特异性DNA结合功能。有关PACT基因及其产物的生物学功能仍有待更多的研究探索。
Osada等[10]在1998年克隆到一个新基因命名为p51。其后又发现几个与p53高度同源的新基因如p73A、p73B、p51A、p51B、p63等。这些基因与p53除了结构上高度同源外,在功能上也存在非常复杂的联系,所以把这一类基因称作为p53基因家族,其生物学功能和意义以及在肿瘤发生中的作用正在研究之中。
四、p53与DNA肿瘤病毒蛋白的相互作用
p53基因产物为核内磷酸化蛋白,具有蛋白质-蛋白质和DNA-蛋白质结合的功能。p53蛋白不仅可与双链DNA结合,也具有与单链DNA结合的能力,除了能与细胞内某些基因产物蛋白结合,还可与一些肿瘤病毒转化蛋白结合并使p53功能失活。 研究认为除p53突变外,肿瘤病毒蛋白与p53结合是p53改变与肿瘤发生关系中最主要的原因。目前已报道的能与p53结合的细胞内蛋白有CBF、E6-AP、ERCC3、HSP70、mdm2、RPA、SP1、TAFⅡ40、TAFⅡ60、TBP和WT1等;能与p53结合的DNA肿瘤病毒蛋白有腺病毒E1B、EBNA5、HPV-E6、SV40T和HBxAg。p53基因与肿瘤病毒相互作用的生物学机制如图2所示。
, http://www.100md.com
注:方框内为最终结果
图2 p53与肿瘤病毒蛋白相互作用与
肿瘤发生机制(参考文献[5])
p53蛋白与SV40 T、腺病毒E1B、EBNA5和HPV6的相互关系及在肿瘤发生中的作用,已有许多报道,现介绍乙肝病毒X蛋白与p53的关系及其在肝癌发生中的作用机制。
乙型肝炎病毒(HBV)为一种小型的DNA病毒,HBV慢性感染者发生肝癌的危险性比无感染者高200倍左右。虽经多年研究,HBV致肝癌的确切机制仍不明确。HBV X基因为4个开放阅读框架中最小的1个,其编码产物HBxAg具有反式激活作用,与肝癌的发生具有密切关系。肝癌中p53突变频率是肿瘤基因突变较高的一种。在一些与HBV感染相关的肝癌中,发现存在p53蛋白细胞内积聚,但p53基因突变率则较低,提示HBxAg可能具有与其他DNA肿瘤病毒编码产物类似的作用,即与p53之间存在蛋白-蛋白结合的关系。为证实这一假设,我们[11,12]应用免疫沉淀和细胞内掺入的方法,于1993年报道了HBxAg与p53存在蛋白-蛋白结合。继之其他实验室进一步研究证实,HBV是继SV40、腺病毒、EBV和HPV之后,第5个病毒编码产物能与p53蛋白结合的DNA病毒。这一现象不仅存在人HBV,也存在于WHV[13]。进一步对HBV基因组进行分析发现,在HBV增强子Ⅰ(enhancer Ⅰ)上游的1047~1059 bp区域,存在经典的p53基因DNA-蛋白质结合位点类似的基因序列,即2个TGCCT正向重复序列。经凝胶电泳迁移率等实验证实,这一序列可特异性地与p53蛋白结合[14]。以上研究初步表明,在HBV感染相关性肝癌中,HBV与p53之间不仅存在蛋白-蛋白(HBxAg-p53)的结合,还存在DNA-蛋白质(HBV-p53)的结合,导致p53蛋白在细胞内积聚,正常的负调节功能丧失。p53在HBV相关性肝癌发生中的可能作用机制如图3所示。
, http://www.100md.com
注:方框内指病毒以外的相关因素
图3 p53与HBV相关性肝癌发生的可能机制
综上所述,关于抑癌基因p53与肿瘤关系,近年来较注重于p53功能、基因间调节、与其他因子相互关系等方面的研究。随着研究的不断深入,进一步加深对p53基因生物学功能和在肿瘤发生发展过程中作用的理解,将更有助于肿瘤的诊断、治疗和预后评估。
参考文献:
[1]Koshland DE Jr. Molecular of the year .Science,1993,262:1953.
[2]El-Deiry WS, Tokino T, Velculescu VE, et al. WAF1, a potential mediator of p53 tumor suppression. Cell ,1993,75:817-825.
, 百拇医药
[3]Bunz F, Dutriaux A, Lengauer C, et al. Requirement for p53 and p21 to sustain G2 arrest after DNA damage. Science,1998,282:1497-1501.
[4]Kamijo T, Weber JD ,Zambetti G, et al. Functioal and physical interactions of the ARF tumor suppressor with p53 and mdm2. Proc Natl Acad Sci USA,1998,95:8292-8297.
[5]Velculescu VE, EL-Deiry WS. Biological and clinical importance of the p53 tumor suppressor gene. Clin Chem, 1996,42:858-869.
, 百拇医药
[6]Schmale H, Bamberger C. A novel protein with strong homology to the tumor suppressor p53. Oncogene ,1997,15:1363-1367.
[7]Nishimori H, Shiratsuchi T, Urano T, et al. A novel brain-specific p53-target gene,BAI1,containing thrombospondin type 1 repeats inhibits experimental angiogenosis. Oncogene,1997,15:2145-2150.
[8]Israeli D, Tessler E, Haupt Y, et al. A novel p53-inducible gene,PAG608, encodes a nuclear zinc finger protein whose overexpression promotes apoptosis. EMBO J,1997,16:4384-4392.
, 百拇医药
[9]Simons A, Melamed-Bessudo C,Wolkowicz R,et al. PACT:cloning and characterization of cellular p53 binding protein that interacts with Rb. Oncogene,1997,14:145-155.
[10]Kaelin WJ. The emerging p53 family. J Natl Cancer Inst,1999,91:594- 598.
[11]朱明华,Feitelgon MA, London WT. HBxAg和p53蛋白结合在原发性肝癌发 生中的意义.中华医学杂志,1993,73:325-327.
[12]Feitelson MA, Zhu MH, Duan LX ,et al. Hepatitis B X antigen and p53 are associated in vitro and in liver tissues from patients with primary hepatocellular Carcinoma. Oncogene, 1993,8:1109-1117.
, 百拇医药
[13]Feitelson MA,Ranganathan PN,Clayton MM, et al. Partial characterization of woodchuck tumor suppressor ,p53, and its interaction with woodchuck hepatitis virus X antigen in hepatocarcinogenesis. Oncogene, 1997,15:327-336.
[14]朱明华,Duan LX,Feitelson MA, et al. 乙型肝炎病毒与肿瘤抑制基因p53 相互作用的生物学功能及意义.病毒学报,1998,14:10-15.
收稿日期:1999-06-21, 百拇医药