EGb761对抗海人藻酸神经毒性的作用
作者:朱俐 高静 张祖暄
单位:朱俐(南通医学院航海医学研究所,南通226001);高静 张祖暄(南京大学医学院,南京210093)
关键词:培养神经元;神经毒性;海人藻酸;EGb761;内酯B
南通医学院学报000105[摘 要] 目的:探讨银杏叶提取物(EGb761)对海人藻酸(KA)引起皮层神经元毒性作用的影响及可能的作用机制。方法:在小鼠原代培养的皮层神经元KA毒性模型上,采用形态学观察,MTT比色分析,ELISA及单细胞内游离钙离子浓度测定等方法。结果:EGb761( 62.5μg/ml)及内酯B(37.5μg/ml)预处理24h可不同程度地对抗KA (0.5mmol/L) 短时间作用引起神经元损伤。结论:EGb761可部分对抗海人藻酸对培养神经元的毒性作用,其作用机制可能与EGb761及其活性成分内酯B维持细胞内钙离子稳态有关。
, http://www.100md.com
[中图分类号] R338.1 [文献标识码] A
[文章编号]1000-2057(2000)01-0013-03
PARTIAL PROTECTIVE EFFECTS OF EGB761 AGAINST KAINATE NEUROTOXICITY
ZHU li
(Institute of Nautical Medicine, Nantong Medical College, Nantong 226001)
GAO Jing ZHANG Zuxuan
(School of Medicine, Nanjing University, Nanjing 210093)
, 百拇医药
[Abstract] Objective:To determine whether the extract of leaves of Ginkgo biloba L ( EGb761) have protective effects against kainate neurotoxicity and what is the possible mechanism.Methods: Morphological observation, MTT staining, ELISA and the intracellular free calcium concentration of single neuron determination were used on kainate neurotoxicity of primary cultures from mouse cerebral cortex. Results: Pretreatment with EGb761(62.5μg/ml)and ginkgolide B (37.5μg/ml)for 24h protected the neurons against KA (0.5 mmol/L)-induced injury in different degree.Conclusions: EGb761 and its active component ginkgolide B partially prevents neurons from kainate neurotoxicity through maintaining the intracellular calcium homeostasis.
, 百拇医药
[Key words] cultured neurons;neurotoxicity;kainaic acid;EGb761;ginkgolide B
兴奋性氨基酸如谷氨酸是哺乳动物脑内含量最高的氨基酸。谷氨酸受体广泛存在于哺乳动物的脑中,根据受体激动剂的特性,形成至少5种兴奋性氨基酸受体类型。自60年代末Olney报道谷氨酸( Glutamate, Glu )有神经毒性作用以来, 兴奋性氨基酸类似物和拮抗剂不断涌现, 从而大大促进了兴奋性氨基酸的研究。 所谓“兴奋性神经毒素”, 最初指的是Glu和海人藻酸( Kainic acid, KA )及其结构类似物可引起哺乳动物特定脑区神经元损伤, 使受损部位的细胞结构发生特征性改变。细胞受损过程中胞外钙离子不断内流、胞内钙库释放钙, 干扰神经元的调节机制, 导致离子、渗透压和电化学的改变,继而造成胞内钙超载和神经元最终死亡。细胞内钙超载可能是兴奋性毒素引起细胞死亡的病理生理学机制[1,2]。
, 百拇医药
银杏叶提取物( Extract of leaves of Ginkgo biloba L.EGb761)对Glu引起的神经元损伤具有一定的神经保护作用[3,4]。EGb761是从银杏科植物叶子经多步骤分离、纯化获得的一种混合物,成分十分复杂, 其主要有效成分为黄酮糖苷类(24%)和萜烯内酯类(6%)。现已初步搞清, 前者具有氧自由基清除功效,后者的内酯B则是血小板活化因子受体的专一性拮抗剂, 具有较高的生物活性[5]。目前国内外已有EGb761胶丸或片剂应市, 用来防治心脑血管系统疾病,但EGb761的药理作用远未搞清。药代动力学研究表明, 至少有60%的EGb761可被机体吸收, 同位素标记物在体内呈广泛而不均匀的分布, 其中脑组织( 皮层、海马和下丘脑 )亦有较高的标记物积聚, 提示EGb761对神经元可能有更直接的保护作用[5]。本实验利用原代培养皮层神经元的KA毒性模型,观察EGb761及内酯B对神经元损伤的影响,同时从单个神经元胞内钙离子浓度变化探讨EGb761可能的作用机制。
, http://www.100md.com
1 材料和方法
1.1 小鼠皮层神经细胞原代培养及KA的毒性作用 参考Choi的方法[6],取孕15~17d的昆明种小鼠胎鼠,无菌条件下迅速分离其大脑皮层,D-hank's平衡盐溶液冲洗2~3遍,尽量剔除血管,剪碎后用终浓度0.125%胰蛋白酶置于37℃下孵育10min,2500rpm离心5min,弃上清,沉淀用Hank's液混匀后再离心。如此反复2次以洗净胰酶,用含10%小牛血清和10%马血清的DMEM培养基制成细胞悬液,台盼蓝染色检测细胞存活率。以1~2×106个细胞/ml的密度接种于预先放有L-多聚赖氨酸的96孔培养板(100μl/孔,用于MTT比色和ELISA测定)、4孔培养板(400μl/孔,孔内置盖玻片供扫描电镜和细胞内钙测定 ), 置37℃、5%CO2培养箱,4h左右贴壁。培养3~5d后加入10-5mmol/L盐酸阿糖胞苷作用1~2d以抑制非神经细胞的过度繁殖,每隔3~4d换一次新鲜培养基,每天以倒置显微镜观察其生长情况。培养14d(14DIV)的皮层细胞吸去培养液,用D-Hank's液洗2遍。换上无血清DMEM培养基,加入不同剂量的KA,37℃下作用一定的时间后,再用Hank's液洗两遍,换上无血清DMEM培养基继续培养24h,相差显微镜下进行形态学观察,用MTT法测定各孔细胞活性或ELISA法测定B-50蛋白免疫活性。
, http://www.100md.com
1.2 MTT比色分析 MTT比色原理是利用活细胞线粒体脱氢酶能将MTT盐还原成蓝紫色的甲瓒颗粒,以颗粒溶解后呈现的颜色深浅反映细胞活性[7]。96孔板每孔加入25μl MTT溶液(终浓度1mg/ml ),37℃下继续培养4h,每孔再加入100μl 20%SDS(溶于50%DMF),37℃放置20h,用微孔酶标仪在570 nm处测定其光吸收值(OD值)。
1.3 ELISA测定神经细胞B-50 蛋白免疫活性 96孔培养板每孔加入100μl 4%多聚甲醛,室温下固定1h,吸去上清,甲醇作用20min(-20℃),PBS荡洗3次,每次5min。吸干残留液后用含10%小牛血清的PBS室温下孵育1h,加入1∶1000的兔抗鼠B-50蛋白抗体(一抗),4℃下放置18h。吸去上清,PBS同上法洗,再加入1∶750的羊抗兔抗体(二抗),37℃下孵育1h。PBS同上法洗,加入底物反应液,37℃下孵育20min后加入2N H2SO4终止反应,以微孔酶标仪于490nm波长处测定其光吸收值。
, 百拇医药
1.4 扫描电镜观察 吸去四孔板中培养液,每孔加4%戊二醛400μl固定1.5h,PBS洗3次,每次3min。再用1%锇酸固定2h,PBS洗3次,每次3mim。后经乙腈逐级脱水,每次20min, 弃去100%乙腈后,再加入100%乙腈,150μl/孔,真空干燥1.5h。取出玻片,金属镀膜仪喷镀金属膜,扫描电镜观察。
1.5 培养神经元胞内[Ca2+]i 的测定 小鼠皮层神经元接种于铺有多聚赖氨酸的盖玻片上,置于四孔板内(直径12mm)。每孔含400μl DMEM培养基,培养3d后用EGb761或内酯B预处理24h, 换上无血清培养基后加KA作用30min。再用Fura 2-AM(2μmol/L )在37℃下孵育45min,用Hank's液洗净, 即时给药组于临测前给予KA。3000r/min离心5min去除胞外的Fura-2,100μl细胞悬液在37℃恒温下用Spex-AR-CM-MIC阳离子检测系统测定单个神经元双波长荧光比值的变化(λex=340和380nm,λem=505nm),以下式计算单细胞内[Ca2+]i:[Ca2+]i=Kd×(FD/Fs)×[(R-Rmin)/(Rmax-R)]。
, http://www.100md.com
2 结 果
2.1 KA对培养神经元的毒性作用 培养14d的小鼠皮层神经元, 用不同浓度KA(0.125~0.5mmol/L)作用30min, 神经元的存活率随KA浓度的增大而下降。0.5mmol/L 的KA分别作用于上述神经元0.5h和24h,0.5h即产生明显的损伤,24h神经元活性下降更明显(图1)。
图1 培养的成熟皮层神经元KA神经毒性作用的剂量和时间依赖关系
A.培养14天的神经细胞用不同浓度的KA(0.125~0.5mmol/L)处理30min;B.培养14天的神经细胞用0.5mmol/L的KA分别处理0.5和24h;平均值±标准差,n=10~12,cP<0.01与0h组相比,fP<0.01与0.5h组相比。
, http://www.100md.com
2.2 EGb761及Gin B对抗KA的毒性作用
2.2.1 形态学变化 培养7d的皮层神经元,胞体透亮,突起长,细胞间建立较好的联系。扫描电镜下可见神经细胞表面光滑,突起完整。KA(0.5mmol/L)作用30min,神经细胞变小,突起断裂。EGb761(62.5μg/ml)预处理24h后再经KA作用,大部分神经元仍保持原有的形态(图2)。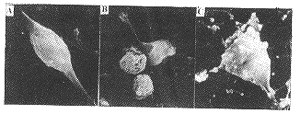
图2 培养7天的皮层神经元经KA和EGb761处理后细胞表面结构的变化
A.未处理的神经元,胞体圆,表面光滑,有正常的突起;B.神经元经0.5mmol/L KA作用30min,显示胞体皱缩和突起丢失;C.神经元经EGb761(62.5μg/ml预处理24h后再用0.5mmol/L作用30min,胞体正常。(×5000)
, 百拇医药
2.2.2 对培养神经元存活的影响 培养14d的皮层神经元经EGb761(2.5μg/ml)预处理24h后,加KA(0.5mmol/L )作用30min, 神经元活性较KA损伤组明显升高, 但并未达对照组水平;当增加EGb761浓度, 对KA毒性作用的对抗不随之增加;当延长KA作用时间达24h,EGb761(62.5μg/ml)不能对抗KA的毒性损伤(图3)。而神经元经EGb761(62.5μg/ml)或 Gin B(37.5μg/ml)预处理24h, 加入 0.5mmol/L KA作用1h,EGb761和Gin B均能明显增加神经元活性,EGb761可完全、Gin B可部分对抗KA毒性作用(图4)。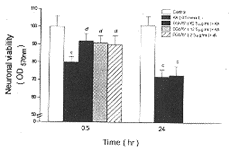
图3 EGb761对KA诱导的小鼠皮层神经元毒性作用的影响
培养14天的神经元经EGb761(2.5~62.5μg/ml)预处理24h,再经KA(0.5mmol/L)作用0.5或24h,KA处理1天用MTT法测定神经元活性。平均值±标准差,n=10~12,cP<0.01与对照组相比,fP<0.01与KA处理组相比。
, 百拇医药
2.2.3 对B-50蛋白免疫活性变化的影响 KA对神经元的损伤也可以通过ELASA方法测定B-50蛋白的免疫活性来反映。离体培养7d的皮层神经元加入0.5mmol/L KA作用30min, B-50蛋白免疫活性显著下降。若用EGb761(62.5μg/ml)或Gin B (37.5μg/ml)预处理24h, B-50蛋白免疫活性有回升的趋势,但差异无显著性。EGb761对抗KA的神经毒性作用似乎与B-50蛋白关系不大(图5)。
图4 EGb761和GinB对KA诱导的B-50蛋白免疫活性下降的影响
培养7天的神经元经EGb761(62.5μg/ml)或GinB(37.5μg/ml)预处理24h,再经KA(0.5mmol/L)作用1h,KA处理后1天用MTT法测定神经元活性。平均值±标准差,n=8,aP>0.05,cP<0.01与0对照组相比,eP<0.05,fP<0.01与KA处理组相比。
, 百拇医药
图5 EGb761和GinB对KA诱导的B-50蛋白免疫活性下降的影响
培养7天的神经元经EGb761(62.5μg/ml)或GinB(37.5μg/ml)预处理24h,再经KA(0.5mmol/L)作用0.5h,KA处理后1天用ELISA法测定神经元B-50蛋白免疫活性。平均值±标准差,n=12,bP<0.05与对照组相比,dP>0.05与KA处理组相比。
2.2.4 对培养神经元胞内游离钙浓度的影响 生长于盖玻片表面的皮层神经元(5DIV)细胞内游离钙浓度([Ca2+]i)平均为101±17nmol/L,加入KA (0.5mmol/L)后[Ca2+]i显著上升。KA(0.5mmol/L)作用30min,[Ca2+]i依然保持较高的水平。但若用EGb761或Gin B预处理24h, 则KA(0.5mmol/L)作用30min后[Ca2+]i没有出现上述显著变化,见表1。 表1 EGb761和Gin B对KA(0.5mmol/L)
, http://www.100md.com
诱导的小鼠皮层神经元[Ca2+]i升高的影响
Group
[Ca2+]i(nmol/L)
n
Control
KAa
KAb
EGb761+KAb
Gin B+KAb
101±17
, 百拇医药
227±84c
237±80c
105±15f
83±9f
20
10
10
10
10
3 讨 论
EGb761具有广泛的生理活性,如抗缺血、抗水肿、抗低氧、清除自由基等。其药理作用也是“多价的”(Polyvalent),主要包括两大方面:一是血管和组织的保护作用,二是对认知的提高作用。近年的研究表明,EG6761及部分活性成份对培养神经元有直接保护作用[8]。本实验结果表明,KA可引起培养的皮层神经元胞体皱缩,突起断裂,神经元活性及B-50蛋白免疫活性显著下降。且神经元活性的下降与KA的浓度和作用时间存在一定的依赖关系。EGb761预处理24h可明显升高神经元的活性, 但是当延长KA的作用时间达24h,也即增加KA的毒性损伤作用时,EGb761 的预保护作用则变得不明显。另一方面,EGb761的主要活性成分之一内酯B也能部分对抗KA的神经毒性作用。但无论是EGb761还是内酯B的抗KA神经毒性作用似乎都与B-50蛋白关系不大。B-50蛋白是一种神经元专一性生长相关蛋白,主要分布于生长锥、轴突及胞体质膜。可作为轴突受损后再生的标记蛋白[9],故推测EGb761的抗KA神经毒性作用是不完全的。
, 百拇医药
以往的工作表明EGb761能保护神经元对抗Glu的损伤,阻止Glu诱发的胞内钙离子浓度的增高[4]。由于谷氨酸受体可大别为两类,即N-甲基-D- 天冬氨酸型(N-methyl-D-aspartate, NMDA)和非NMDA型。一般认为KA作为谷氨酸受体的激动剂之一,主要作用于非NMDA型谷氨酸受体,引起钠、钾离子内流和膜的去极化。 近年的研究发现, KA的兴奋性神经毒性作用也与胞内钙离子水平的提高密切相关[10]。本实验结果则提示KA可提高细胞内游离钙浓度平均水平,而EGb761或Gin B 预处理24h,神经元胞内钙离子浓度可保持相对的稳定。由此可见,EGb761对抗KA的神经毒性作用可能与其维持细胞钙离子稳态有关,而内酯B可能起了主要作用。
[参考文献]
[1] Gwag BJ,Canzoniero LM,Sensi SL,Demaro JA,Koh JY,Goldberg MP,Jacquin M,Choi DW.Calcium ionophores can induce either apoptosis or necrosis in cultured cortical neurons[J].Neuroscience,1999,90(4)∶1339.
, 百拇医药
[2] Mattson MP.Calcium and free radicals: Mediators of neurotrophic factor and excitatory transmitter-regulated developmental plasticity and cell death[J].Perspect Dev Neurobiol,1996,3(2)∶79.
[3] Lin L,Gu HM, Gao J,et al.Protective effects of ginkgotins on glutamate neurotoxicity in vitro[J].J Nanjing Univ,(Natural Science Edition 1995),31(3)∶514.
[4] Zhu L, Wu J,Liao H,et al.Antagonistic effects of extract from leaves of Ginkgo biloba on glutamate neurotoxicity[J].Acta pharmacologica Sinica,1997,18(4)∶344.
, 百拇医药
[5] DeFeudis FV.Ginkgo biloba extract ( EGb761 ): pharmacological activities and clinical applications[M].Pris: Elsevir,1991,9~94.
[6] Choi DW, Maulucci-Gedde M, Kriegstein AR.Glutamate neurotoxicity in cortical cell culture[J].J Neurosci,1987,7(2)∶357.
[7] Hansen MB, Nielsen SE, Berg K.Re-examination and further development of a precise and rapid dye method for measuring cell growth/cell kill[J].J Immunol Methods, 1989,119∶203.
, http://www.100md.com
[8] Ahlemeyer B,Mowes A,Krieglstein J.Inhibition of serum deprivation and staurosporine-induced neuronal apoptosis by Ginkgo biloba extract and some of its constituents[J].Eur J Pharmacol,1999,367(2~3)∶423.
[9] Campagne MVL, Dotti CG, Verkleij AJ, Gispen WH and Oestreicher AB.B-50/GAP43 localization on membranes of putative transport vesicles in the cell body, neurites and growth cones of cultured hippocampal neurons[J].Neurosci Lett,1992,137∶129.
[10] Leski ML, Valentine SL, Coyle JT.L-type voltage-gated calcium channels modulate kainic acid neurotoxicity in cerebellar granule cells[J].Brain Res,1999, 828(1~2)∶27.
(1999-11-15收稿), http://www.100md.com
单位:朱俐(南通医学院航海医学研究所,南通226001);高静 张祖暄(南京大学医学院,南京210093)
关键词:培养神经元;神经毒性;海人藻酸;EGb761;内酯B
南通医学院学报000105[摘 要] 目的:探讨银杏叶提取物(EGb761)对海人藻酸(KA)引起皮层神经元毒性作用的影响及可能的作用机制。方法:在小鼠原代培养的皮层神经元KA毒性模型上,采用形态学观察,MTT比色分析,ELISA及单细胞内游离钙离子浓度测定等方法。结果:EGb761( 62.5μg/ml)及内酯B(37.5μg/ml)预处理24h可不同程度地对抗KA (0.5mmol/L) 短时间作用引起神经元损伤。结论:EGb761可部分对抗海人藻酸对培养神经元的毒性作用,其作用机制可能与EGb761及其活性成分内酯B维持细胞内钙离子稳态有关。
, http://www.100md.com
[中图分类号] R338.1 [文献标识码] A
[文章编号]1000-2057(2000)01-0013-03
PARTIAL PROTECTIVE EFFECTS OF EGB761 AGAINST KAINATE NEUROTOXICITY
ZHU li
(Institute of Nautical Medicine, Nantong Medical College, Nantong 226001)
GAO Jing ZHANG Zuxuan
(School of Medicine, Nanjing University, Nanjing 210093)
, 百拇医药
[Abstract] Objective:To determine whether the extract of leaves of Ginkgo biloba L ( EGb761) have protective effects against kainate neurotoxicity and what is the possible mechanism.Methods: Morphological observation, MTT staining, ELISA and the intracellular free calcium concentration of single neuron determination were used on kainate neurotoxicity of primary cultures from mouse cerebral cortex. Results: Pretreatment with EGb761(62.5μg/ml)and ginkgolide B (37.5μg/ml)for 24h protected the neurons against KA (0.5 mmol/L)-induced injury in different degree.Conclusions: EGb761 and its active component ginkgolide B partially prevents neurons from kainate neurotoxicity through maintaining the intracellular calcium homeostasis.
, 百拇医药
[Key words] cultured neurons;neurotoxicity;kainaic acid;EGb761;ginkgolide B
兴奋性氨基酸如谷氨酸是哺乳动物脑内含量最高的氨基酸。谷氨酸受体广泛存在于哺乳动物的脑中,根据受体激动剂的特性,形成至少5种兴奋性氨基酸受体类型。自60年代末Olney报道谷氨酸( Glutamate, Glu )有神经毒性作用以来, 兴奋性氨基酸类似物和拮抗剂不断涌现, 从而大大促进了兴奋性氨基酸的研究。 所谓“兴奋性神经毒素”, 最初指的是Glu和海人藻酸( Kainic acid, KA )及其结构类似物可引起哺乳动物特定脑区神经元损伤, 使受损部位的细胞结构发生特征性改变。细胞受损过程中胞外钙离子不断内流、胞内钙库释放钙, 干扰神经元的调节机制, 导致离子、渗透压和电化学的改变,继而造成胞内钙超载和神经元最终死亡。细胞内钙超载可能是兴奋性毒素引起细胞死亡的病理生理学机制[1,2]。
, 百拇医药
银杏叶提取物( Extract of leaves of Ginkgo biloba L.EGb761)对Glu引起的神经元损伤具有一定的神经保护作用[3,4]。EGb761是从银杏科植物叶子经多步骤分离、纯化获得的一种混合物,成分十分复杂, 其主要有效成分为黄酮糖苷类(24%)和萜烯内酯类(6%)。现已初步搞清, 前者具有氧自由基清除功效,后者的内酯B则是血小板活化因子受体的专一性拮抗剂, 具有较高的生物活性[5]。目前国内外已有EGb761胶丸或片剂应市, 用来防治心脑血管系统疾病,但EGb761的药理作用远未搞清。药代动力学研究表明, 至少有60%的EGb761可被机体吸收, 同位素标记物在体内呈广泛而不均匀的分布, 其中脑组织( 皮层、海马和下丘脑 )亦有较高的标记物积聚, 提示EGb761对神经元可能有更直接的保护作用[5]。本实验利用原代培养皮层神经元的KA毒性模型,观察EGb761及内酯B对神经元损伤的影响,同时从单个神经元胞内钙离子浓度变化探讨EGb761可能的作用机制。
, http://www.100md.com
1 材料和方法
1.1 小鼠皮层神经细胞原代培养及KA的毒性作用 参考Choi的方法[6],取孕15~17d的昆明种小鼠胎鼠,无菌条件下迅速分离其大脑皮层,D-hank's平衡盐溶液冲洗2~3遍,尽量剔除血管,剪碎后用终浓度0.125%胰蛋白酶置于37℃下孵育10min,2500rpm离心5min,弃上清,沉淀用Hank's液混匀后再离心。如此反复2次以洗净胰酶,用含10%小牛血清和10%马血清的DMEM培养基制成细胞悬液,台盼蓝染色检测细胞存活率。以1~2×106个细胞/ml的密度接种于预先放有L-多聚赖氨酸的96孔培养板(100μl/孔,用于MTT比色和ELISA测定)、4孔培养板(400μl/孔,孔内置盖玻片供扫描电镜和细胞内钙测定 ), 置37℃、5%CO2培养箱,4h左右贴壁。培养3~5d后加入10-5mmol/L盐酸阿糖胞苷作用1~2d以抑制非神经细胞的过度繁殖,每隔3~4d换一次新鲜培养基,每天以倒置显微镜观察其生长情况。培养14d(14DIV)的皮层细胞吸去培养液,用D-Hank's液洗2遍。换上无血清DMEM培养基,加入不同剂量的KA,37℃下作用一定的时间后,再用Hank's液洗两遍,换上无血清DMEM培养基继续培养24h,相差显微镜下进行形态学观察,用MTT法测定各孔细胞活性或ELISA法测定B-50蛋白免疫活性。
, http://www.100md.com
1.2 MTT比色分析 MTT比色原理是利用活细胞线粒体脱氢酶能将MTT盐还原成蓝紫色的甲瓒颗粒,以颗粒溶解后呈现的颜色深浅反映细胞活性[7]。96孔板每孔加入25μl MTT溶液(终浓度1mg/ml ),37℃下继续培养4h,每孔再加入100μl 20%SDS(溶于50%DMF),37℃放置20h,用微孔酶标仪在570 nm处测定其光吸收值(OD值)。
1.3 ELISA测定神经细胞B-50 蛋白免疫活性 96孔培养板每孔加入100μl 4%多聚甲醛,室温下固定1h,吸去上清,甲醇作用20min(-20℃),PBS荡洗3次,每次5min。吸干残留液后用含10%小牛血清的PBS室温下孵育1h,加入1∶1000的兔抗鼠B-50蛋白抗体(一抗),4℃下放置18h。吸去上清,PBS同上法洗,再加入1∶750的羊抗兔抗体(二抗),37℃下孵育1h。PBS同上法洗,加入底物反应液,37℃下孵育20min后加入2N H2SO4终止反应,以微孔酶标仪于490nm波长处测定其光吸收值。
, 百拇医药
1.4 扫描电镜观察 吸去四孔板中培养液,每孔加4%戊二醛400μl固定1.5h,PBS洗3次,每次3min。再用1%锇酸固定2h,PBS洗3次,每次3mim。后经乙腈逐级脱水,每次20min, 弃去100%乙腈后,再加入100%乙腈,150μl/孔,真空干燥1.5h。取出玻片,金属镀膜仪喷镀金属膜,扫描电镜观察。
1.5 培养神经元胞内[Ca2+]i 的测定 小鼠皮层神经元接种于铺有多聚赖氨酸的盖玻片上,置于四孔板内(直径12mm)。每孔含400μl DMEM培养基,培养3d后用EGb761或内酯B预处理24h, 换上无血清培养基后加KA作用30min。再用Fura 2-AM(2μmol/L )在37℃下孵育45min,用Hank's液洗净, 即时给药组于临测前给予KA。3000r/min离心5min去除胞外的Fura-2,100μl细胞悬液在37℃恒温下用Spex-AR-CM-MIC阳离子检测系统测定单个神经元双波长荧光比值的变化(λex=340和380nm,λem=505nm),以下式计算单细胞内[Ca2+]i:[Ca2+]i=Kd×(FD/Fs)×[(R-Rmin)/(Rmax-R)]。
, http://www.100md.com
2 结 果
2.1 KA对培养神经元的毒性作用 培养14d的小鼠皮层神经元, 用不同浓度KA(0.125~0.5mmol/L)作用30min, 神经元的存活率随KA浓度的增大而下降。0.5mmol/L 的KA分别作用于上述神经元0.5h和24h,0.5h即产生明显的损伤,24h神经元活性下降更明显(图1)。
图1 培养的成熟皮层神经元KA神经毒性作用的剂量和时间依赖关系
A.培养14天的神经细胞用不同浓度的KA(0.125~0.5mmol/L)处理30min;B.培养14天的神经细胞用0.5mmol/L的KA分别处理0.5和24h;平均值±标准差,n=10~12,cP<0.01与0h组相比,fP<0.01与0.5h组相比。
, http://www.100md.com
2.2 EGb761及Gin B对抗KA的毒性作用
2.2.1 形态学变化 培养7d的皮层神经元,胞体透亮,突起长,细胞间建立较好的联系。扫描电镜下可见神经细胞表面光滑,突起完整。KA(0.5mmol/L)作用30min,神经细胞变小,突起断裂。EGb761(62.5μg/ml)预处理24h后再经KA作用,大部分神经元仍保持原有的形态(图2)。
图2 培养7天的皮层神经元经KA和EGb761处理后细胞表面结构的变化
A.未处理的神经元,胞体圆,表面光滑,有正常的突起;B.神经元经0.5mmol/L KA作用30min,显示胞体皱缩和突起丢失;C.神经元经EGb761(62.5μg/ml预处理24h后再用0.5mmol/L作用30min,胞体正常。(×5000)
, 百拇医药
2.2.2 对培养神经元存活的影响 培养14d的皮层神经元经EGb761(2.5μg/ml)预处理24h后,加KA(0.5mmol/L )作用30min, 神经元活性较KA损伤组明显升高, 但并未达对照组水平;当增加EGb761浓度, 对KA毒性作用的对抗不随之增加;当延长KA作用时间达24h,EGb761(62.5μg/ml)不能对抗KA的毒性损伤(图3)。而神经元经EGb761(62.5μg/ml)或 Gin B(37.5μg/ml)预处理24h, 加入 0.5mmol/L KA作用1h,EGb761和Gin B均能明显增加神经元活性,EGb761可完全、Gin B可部分对抗KA毒性作用(图4)。
图3 EGb761对KA诱导的小鼠皮层神经元毒性作用的影响
培养14天的神经元经EGb761(2.5~62.5μg/ml)预处理24h,再经KA(0.5mmol/L)作用0.5或24h,KA处理1天用MTT法测定神经元活性。平均值±标准差,n=10~12,cP<0.01与对照组相比,fP<0.01与KA处理组相比。
, 百拇医药
2.2.3 对B-50蛋白免疫活性变化的影响 KA对神经元的损伤也可以通过ELASA方法测定B-50蛋白的免疫活性来反映。离体培养7d的皮层神经元加入0.5mmol/L KA作用30min, B-50蛋白免疫活性显著下降。若用EGb761(62.5μg/ml)或Gin B (37.5μg/ml)预处理24h, B-50蛋白免疫活性有回升的趋势,但差异无显著性。EGb761对抗KA的神经毒性作用似乎与B-50蛋白关系不大(图5)。
图4 EGb761和GinB对KA诱导的B-50蛋白免疫活性下降的影响
培养7天的神经元经EGb761(62.5μg/ml)或GinB(37.5μg/ml)预处理24h,再经KA(0.5mmol/L)作用1h,KA处理后1天用MTT法测定神经元活性。平均值±标准差,n=8,aP>0.05,cP<0.01与0对照组相比,eP<0.05,fP<0.01与KA处理组相比。
, 百拇医药
图5 EGb761和GinB对KA诱导的B-50蛋白免疫活性下降的影响
培养7天的神经元经EGb761(62.5μg/ml)或GinB(37.5μg/ml)预处理24h,再经KA(0.5mmol/L)作用0.5h,KA处理后1天用ELISA法测定神经元B-50蛋白免疫活性。平均值±标准差,n=12,bP<0.05与对照组相比,dP>0.05与KA处理组相比。
2.2.4 对培养神经元胞内游离钙浓度的影响 生长于盖玻片表面的皮层神经元(5DIV)细胞内游离钙浓度([Ca2+]i)平均为101±17nmol/L,加入KA (0.5mmol/L)后[Ca2+]i显著上升。KA(0.5mmol/L)作用30min,[Ca2+]i依然保持较高的水平。但若用EGb761或Gin B预处理24h, 则KA(0.5mmol/L)作用30min后[Ca2+]i没有出现上述显著变化,见表1。 表1 EGb761和Gin B对KA(0.5mmol/L)
, http://www.100md.com
诱导的小鼠皮层神经元[Ca2+]i升高的影响
Group
[Ca2+]i(nmol/L)
n
Control
KAa
KAb
EGb761+KAb
Gin B+KAb
101±17
, 百拇医药
227±84c
237±80c
105±15f
83±9f
20
10
10
10
10
3 讨 论
EGb761具有广泛的生理活性,如抗缺血、抗水肿、抗低氧、清除自由基等。其药理作用也是“多价的”(Polyvalent),主要包括两大方面:一是血管和组织的保护作用,二是对认知的提高作用。近年的研究表明,EG6761及部分活性成份对培养神经元有直接保护作用[8]。本实验结果表明,KA可引起培养的皮层神经元胞体皱缩,突起断裂,神经元活性及B-50蛋白免疫活性显著下降。且神经元活性的下降与KA的浓度和作用时间存在一定的依赖关系。EGb761预处理24h可明显升高神经元的活性, 但是当延长KA的作用时间达24h,也即增加KA的毒性损伤作用时,EGb761 的预保护作用则变得不明显。另一方面,EGb761的主要活性成分之一内酯B也能部分对抗KA的神经毒性作用。但无论是EGb761还是内酯B的抗KA神经毒性作用似乎都与B-50蛋白关系不大。B-50蛋白是一种神经元专一性生长相关蛋白,主要分布于生长锥、轴突及胞体质膜。可作为轴突受损后再生的标记蛋白[9],故推测EGb761的抗KA神经毒性作用是不完全的。
, 百拇医药
以往的工作表明EGb761能保护神经元对抗Glu的损伤,阻止Glu诱发的胞内钙离子浓度的增高[4]。由于谷氨酸受体可大别为两类,即N-甲基-D- 天冬氨酸型(N-methyl-D-aspartate, NMDA)和非NMDA型。一般认为KA作为谷氨酸受体的激动剂之一,主要作用于非NMDA型谷氨酸受体,引起钠、钾离子内流和膜的去极化。 近年的研究发现, KA的兴奋性神经毒性作用也与胞内钙离子水平的提高密切相关[10]。本实验结果则提示KA可提高细胞内游离钙浓度平均水平,而EGb761或Gin B 预处理24h,神经元胞内钙离子浓度可保持相对的稳定。由此可见,EGb761对抗KA的神经毒性作用可能与其维持细胞钙离子稳态有关,而内酯B可能起了主要作用。
[参考文献]
[1] Gwag BJ,Canzoniero LM,Sensi SL,Demaro JA,Koh JY,Goldberg MP,Jacquin M,Choi DW.Calcium ionophores can induce either apoptosis or necrosis in cultured cortical neurons[J].Neuroscience,1999,90(4)∶1339.
, 百拇医药
[2] Mattson MP.Calcium and free radicals: Mediators of neurotrophic factor and excitatory transmitter-regulated developmental plasticity and cell death[J].Perspect Dev Neurobiol,1996,3(2)∶79.
[3] Lin L,Gu HM, Gao J,et al.Protective effects of ginkgotins on glutamate neurotoxicity in vitro[J].J Nanjing Univ,(Natural Science Edition 1995),31(3)∶514.
[4] Zhu L, Wu J,Liao H,et al.Antagonistic effects of extract from leaves of Ginkgo biloba on glutamate neurotoxicity[J].Acta pharmacologica Sinica,1997,18(4)∶344.
, 百拇医药
[5] DeFeudis FV.Ginkgo biloba extract ( EGb761 ): pharmacological activities and clinical applications[M].Pris: Elsevir,1991,9~94.
[6] Choi DW, Maulucci-Gedde M, Kriegstein AR.Glutamate neurotoxicity in cortical cell culture[J].J Neurosci,1987,7(2)∶357.
[7] Hansen MB, Nielsen SE, Berg K.Re-examination and further development of a precise and rapid dye method for measuring cell growth/cell kill[J].J Immunol Methods, 1989,119∶203.
, http://www.100md.com
[8] Ahlemeyer B,Mowes A,Krieglstein J.Inhibition of serum deprivation and staurosporine-induced neuronal apoptosis by Ginkgo biloba extract and some of its constituents[J].Eur J Pharmacol,1999,367(2~3)∶423.
[9] Campagne MVL, Dotti CG, Verkleij AJ, Gispen WH and Oestreicher AB.B-50/GAP43 localization on membranes of putative transport vesicles in the cell body, neurites and growth cones of cultured hippocampal neurons[J].Neurosci Lett,1992,137∶129.
[10] Leski ML, Valentine SL, Coyle JT.L-type voltage-gated calcium channels modulate kainic acid neurotoxicity in cerebellar granule cells[J].Brain Res,1999, 828(1~2)∶27.
(1999-11-15收稿), http://www.100md.com