牛泡沫病毒反式激活因子在内部启动子上应答元件的研究
作者:张莉 乔文涛 刘淑红 王金忠 陈启民 耿运琪
单位:南开大学生命科学学院,天津 300071
关键词:牛泡沫病毒;内部启动子(IP);转录反式激活因子(Tas);反式激活因子应答元件(TRE);TATA盒
病毒学报000309 摘要:泡沫病毒的基因转录依赖至少两个不同的启动子:LTR调节病毒结构蛋白的表达,而内部启动子(IP)则起始调节蛋白mRNA的转录。牛泡沫病毒(BFV)在env与3′LTR之间有两个重叠的开放阅读框架orf-1和orf-2,分别编码BFV ORF-1、ORF-2等多种调节蛋白。这些蛋白中BFV ORF-1为转录激活因子,称为Tas。Tas对LTR及IP均有反式激活作用。BFV中第二类启动子IP的存在反映了泡沫病毒基因调控的复杂性。已从BFV3026中国毒株中克隆了基因组区段8 572~9 509,并通过测序和功能分析证明该区段包含了BFV IP。为了对IP进行更精确的定位,对其进行了进一步的缺失分析,将完整的IP定位在9 117~9 405之间。该启动子的TATA盒位于91 280~9 285,是IP必不可少的元件之一。杂合启动子和缺失分析表明,TATA盒上游160bp的区域内包含了IP的Tas应答元件(TREIP),其功能类似于增强子。Tas对IP具有强烈的激活作用,而C端缺失的ORF-1蛋白或ORF-2蛋白均不足以激活IP的表达。
, 百拇医药
中图分类号:S852.65+9.3; Q75 文献标识码:A
文章编号:1000-8721(2000)03-0232-06
Regulatory Elements That Mediate Transactivation by Tas in the Internal Promoter of Bovine Foamy Virus
ZHANG Li,QIAO Wen-tao,LIU Shu-hong,WANG Jin-zhon,CHEN Qi-min,GENG Yun-qi
(College of Life Sciences, Nankai University, Tianjin 300071, China)
Abstract: Foamy viruses are unusual among retroviruses in that they contain two distinct promoter elements. They are the long terminal repeat (LTR) promoter which regulates expression of the viral structural proteins, and a second internal promoter that locates towards the 3' end of the env gene and directs expression of the viral auxiliary proteins. One of these auxiliary proteins is a potent transcriptional transactivator, termed Tas in BFV. That is critical for foamy virus replication. Previously, we have cloned the intact BFV IP and demonstrated it to be highly responsive to the Tas protein. Here, we extended this earlier work by using deleted mutants and hybrid construct to localize the IP more precisely and to identify and characterize the Tas responsive element (TREIP) . Based on these data, we propose that genomic region 9117~9405nt contains all the sequences required for full Tas responsiveness. The importance of a TATA box for the BFV internal promoter is established by deletion analysis. The cis-acting sequences in the BFV env gene required for response to Tas are contained within a 160-bp element located 5' to the TATA box in the internal promoter. This Tas responsive element in the internal promoter functions in an orientation-independent fashion in a heterologous promoter and thus has the properties of an enhancer which depends on Tas activity. Alignments reveal that the BFV internal promoter and the BFV LTR have little sequence relatedness. In summary, the genome of bovine foamy virus directs viral transcription through both the promoter in the LTR and an internal promoter within the env gene, and each promoter contains unique enhancer-like elements regulated by the viral transactivator. And it seems that the Tas is the only accessory BFV protein necessary for viral replication in vitro.
, 百拇医药
Key words: bovine foamy virus; internal promoter; Tas; Tas responsive element; long terminal repeat promoter
泡沫病毒属于复杂的反转录病毒,它们不仅有复杂的基因组结构,而且具有复杂的基因表达调控机制。泡沫病毒自身编码一个病毒复制所必需的转录反式激活因子Tas。Tas可极大地激活病毒5′LTR起始的基因表达,而且这种激活作用在各种哺乳类乃至禽类细胞中均存在。BFV为非灵长类泡沫病毒,在env和3′LTR之间有两个开放阅读框架(ORF),其中第一个ORF编码BFV Tas蛋白[1]。Tas通过LTR U3区的多个顺式调控位点调节基因表达。本室以前的研究证明除了LTR外,在BFV env基因3′端存在另一个转录启动子,称为内部启动子[2],其活性亦强烈依赖于Tas。IP的存在使泡沫病毒基因表达调节机制比其它反转录病毒更显复杂。所以研究IP是研究泡沫病毒基因调控机制的一个重要环节。为了找到IP的精确位置,同时为了探寻BFV Tas在IP上的靶序列,本文构建了一系列IP缺失质粒以及IP/LTR杂合质粒,将它们转染牛肺细胞系BL-12,引入luc基因作瞬时表达分析。结果将完整的IP定位于基因组9?117~9?405之间。
, 百拇医药
材料与方法
1 病毒Hirt DNA的提取 反转录病毒感染宿主细胞后首先反转录成双链DNA,在反转录病毒感染的细胞的细胞核内常常存在一些非整合的线形或环状双链DNA分子,通常称为Hirt DNA。按文献[3]的方法从被BFV3026病毒株感染的新生牛肺细胞(NBL)中抽提NBL/BFV3026的Hirt DNA,并用TE缓冲液溶解。
2 引物的设计与PCR反应 参考有关HFV[4]和SFV-1[5]文献及最新发表的BFV基因组序列[6],根据env基因中的保守序列设计多对引物(表1),旨在扩增IP的缺失质粒。PCR反应条件为:94℃ 3min,50℃ 1min,72℃ 1min,1个循环;94℃ 30s,50℃ 1min,72℃ 1min,30个循环;94℃ 30s,50℃ 1min,72℃ 10min,1个循环。
, 百拇医药
3 缺失质粒的构建 质粒pBFV-IP-1-luc为本室构建[2],已证明该质粒上的8?572~9?509基因组片段包含了完整的牛泡沫病毒内部启动子。以此质粒为出发质粒,通过PCR方法构建了一系列IP缺失质粒pBFV-IP(2~6)-luc。质粒pBORF-1(1~249aa)、pBORF(1+2)、pBORF2、pBORF1(1~217aa)、pbasic-P由本室构建[7]。pBORF-1(1~249aa)表达完整的BFV ORF-1蛋白,即BFV的反式激活因子Tas;pBORF(1+2)表达orf-1、orf-2所编码的ORF-1、ORF-2、Bet等;pBORF-1(1~217aa)表达ORF-1蛋白的1~217aa肽段;pBFV ORF-2表达完整的BFV ORF-2蛋白(335aa)。这些表达质粒的上游启动子均为强启动子RSV LTR。pbasic-P是将BFV LTR的基本启动子(BFV基因组843~1012区段)克隆在luc基因上游[7]。 质粒pUC19、 pCMV-lacZ、 宿主菌E.coli JM103、TG1均由本室保存。
, 百拇医药
表1 构建质粒采用的引物
Table 1 Primers used in plasmid construction Plasmid
Actual genomic regions
contained in the plasmid
Upper/lower primers
pBFVIP-1-luc
BFV bp 8572~9509
P8564-8592:5′-ATA GGA AGT CGA CTG GAC TGG TCA GTA CT-3′
, http://www.100md.com SalⅠ
P9488-9517:5′-GAT AAG CTT TAT CCC AAT AAT GAC CGG TTT-3
HindⅢ
pBFVIP-2-luc
BFV bp 9053~9509
P9048-90655′ TCA TGT CGA CCC ACA CTG 3′
SalⅠ
p9488-9517:5′-GAT AAG CTT TAT CCC AAT AAT GAC CGG TTT-3′
HindⅢ
, 百拇医药
pBFVIP-3-luc
BFV bp 9053~9405
P9048-9065 5′ TCA TGT CGA C CC ACA CTG 3′
SalⅠ
P9395-9414 5′ CTC AAA GCT TAC GCA GCT AC 3′
HindⅢ
pBFVIP-4-luc
BFV bp 9117~9405
P9112-9130 5′ CTC TGT CGA CTG CTA TGGT 3′
, 百拇医药
SalⅠ
P9395-9414 5′ CTC AAA GCT TAC GCA GCT AC 3′
HindⅢ
pBFVIP-5-luc
BFV bp 9205~9405
P9200-9218 5′ AGA GGT CGA CTT CCT CATT 3′
SalⅠ
P9395-9414 5′ CTC AAA GCT TAC GCA GCT AC 3′
HindⅢ
pBFVIP-6-luc
, http://www.100md.com
BFV bp 9053~9276
P9048-9065 5′ TCA TGT CGA CCC ACA CTG 3′
SalⅠ
P9267-9285 5′ TTT AAA GCT TCC CAA GTGC 3′
HindⅢ
4 酶和试剂 实验用限制性内切酶、E.coli DNA聚合酶Ⅰ(K.F)、T4连接酶购自华美生物工程公司。荧光素酶检测试剂盒购自美国Promega公司。DMEM细胞培养基、青霉素/链霉素(p/s)、hepes及Trypsin购自美国GIBCO/BRL公司。
5 质粒的提取、酶切、DNA片段制备、连接及转化 其操作均参照文献[8]。
, 百拇医药
6 细胞培养和转染 用含10%胎牛血清的DMEM,37℃5%CO2培养。NBL细胞用于病毒繁殖,BL-12细胞系用于转染。转染时,BL-12以2.5×105ml-1接种于6孔板,每孔2ml。转染采用阳离子脂质体法(LipofectAMINE Reagent)。具体转染步骤参见GIBCO/BRL产品说明书。转染除用pUC19作对照质粒外,均设有β-gal内对照系统。ONPG、β-galactosidase购自Sigma公司。2天后裂解细胞测定Luc酶活力。细胞裂解液和Luc酶活力测定试剂盒购自Promega公司。按试剂盒提供的方法,用生物化学发光测量仪(上海检测技术所检测仪器厂生产)测定化学发光强度,方法原理见文献[9]。Luc酶的活力以4S内的光强度表示。结 果
1 完整的IP位于基因组9?117~9?405之间
本室以前的工作已证实了BFV env基因3′末端(基因组8?564~9?517区段)编码一个可被Tas激活的内部启动子[2]。PCR扩增的该区段被插入到无启动子的萤火虫荧光素酶基因上游构成表达质粒pBFV-IP-1-luc。Luc分析表明,单独转染时,pBFV-IP-1-luc只有很低的基础Luc活性,但当与pBORF-1共转染时,活性上升了145倍(图1),这说明IP的活性强烈依赖于Tas。
, http://www.100md.com
为了将IP在基因组上的位置定位得更加精细,我们以质粒pBFV-IP-1-luc为出发质粒,构建了IP的一系列缺失体pBFV-IP(2~6)-luc。将它们分别单独转染或与pBORF-1质粒共转染BL-12细胞系,转染结果如图1所示。克隆片段的相对大小及它们在基因组中的位置在图中亦有表示。由于这些缺失体的基础活性近似,所以推断IP中不含负调控区。当下游保持在9?509,上游缺失481 bp至基因组9?053时(pBFV-IP-2-luc),激活倍数不但没有降低,反而大大提高,由此可以推断缺失掉的这481 bp不影响Tas对IP的激活作用,是无关序列;而当上游为9?053,下游缺失至9?405时(pBFV-IP-3-luc),激活程度并不低于Tas对pBFV-IP-2-luc的作用,所以同样说明下游9?405~9?509这段序列也与IP的功能关系不大。我们此时对上游进行进一步的缺失至9?117(pBFV-IP-4-luc),发现激活倍数达到了350倍。而若继续缺失至9?205时(pBFV-IP-5-luc),激活活性则有所下降。说明IP的上限应定在9?117。在这一基础上,我们对IP的下游又进行了缺失,直至9?276。从图1可看出,激活倍数降到了最低水平3.6,与其它质粒相比可认为没有激活。由此可见,下游9?276~9?405也是IP行使功能必不可少的。因此,我们将IP的下限定在9?405。综上所述,我们可将完整的IP定位于BFV基因组9?117~9?405之间。
, 百拇医药
图1 IP系列缺失体及转染BL-12细胞系的Luc分析结果
Figure 1 A series of IP deleted mutants and the outcome of Luc assay when transfected them into BL-12 cell line
A.Schematic diagram of BFV IP and the amplificated genome regions by PCR;luc.firefly luciferase gene(upper).Lines represent BFV IP regions remained after deletion.B.“-Tas”means cotransfection of the reporter plasmid and pUC19.This value represents basal promoter activity.“+Tas”means cotransfection of the reporter plasmid and the transactivator plasmid (pBFVORF-1).a.Each experiment was done at least 3 times in triplicate and each plate was transfected with 0.2μg of deleted luciferase expression plasmid, 0.75μg of pBFVORF-1,0.6μg of pCMV β-gal and 3 μg of Lipofect AMINE(GIBCO/BRL).The cells were harvested to determine Luc activity at 48h after transfection.Luciferase activities were normalized for β-galactosidase.Generally,less than 15% variation in replicate samples was observed.
, http://www.100md.com
2 Tas对IP的激活作用必须有TATA盒的参与
序列分析和同源比较表明:与BFV IP行使功能有重要关系的9?276~9?405这段区域中含有典型的真核启动子TATA盒。与其它泡沫病毒相比,该TATA盒及其附近的序列十分保守,均为TATAAAAGA*CA*AT。而除TATA盒外,9?276~9?405中未发现其它元件。所以IP的TATA盒是起始转录不可缺少的元件。这个结论同时也证实了IP起始转录是通过RNA pol Ⅱ复合体与TATA盒作用而实现的。
3 BFV IP上的Tas应答元件功能类似于增强子
为了深入研究Borf-1激活IP起始基因表达调控的特点,并且为了探寻Tas在IP上的作用位点,本文对IP TATA盒上游160bp的序列(9?117~9?276)与Borf-1之间的作用特点作了进一步的研究。质粒pbasic-P在luc基因上游含有BFV LTR的基本启动子,但缺少增强子元件,所以这个载体可用于研究增强子[7]。利用此质粒,我们构建了IP-basic P杂合启动子。BFV IP TATA盒近上游160bp的序列被克隆到LTR基本启动子(basic promoter)上游构建杂合体pIPenh-basic-P。瞬时分析实验表明,单独转染杂合体时Luc活性不高。但当它与激活质粒pBORF-1共转染时,Luc活性上升了212倍(图2),而且其激活活性也远远高于pbasic-P与pBORF-1共转染时活性。由此可见:IP TATA盒上游160bp序列强烈依赖于Tas,且有可能以增强子的方式发挥作用。结合杂合启动子以及图1中的缺失分析,我们确认这160bp IP序列中含有Tas作用元件TREIP。
, 百拇医药
4 Borf-1蛋白是IP起始基因表达唯一所需的病毒辅助蛋白
本室以前的工作已证明,BFV LTR起始转录依赖于BFV反式激活因子Tas蛋白,而ORF-2蛋白或C端缺失的ORF-1蛋白均不足以激活BFV LTR的表达[7]。从表2可以看出,这个结论同样适用于IP。本文采用野生型IP质粒pBFV-IP-4-luc作为报告质粒,分别与pBORF-1(1~249aa)、pBORF(1+2)、pBORF2、pBORF1(1~217aa)共转染BL-12,结果见表2。将pBFV-IP-4-luc与无关质粒pUC19共转染时的Luc值设为负对照。当将表达完整Tas的pBORF-1(1~249aa)质粒与pBFV-IP-4-luc共转染时,激活倍数为287;而当pBORF(1+2)与报告质粒共转染时,激活倍数只有8。即使在体系中再加入pBORF-1(1~249aa),激活活性也恢复不到pBORF1(1~249aa)单独与pBFV-IP-4-luc共转染时的水平。说明pBFVORF(1+2)表达产物中有部分蛋白不参与对报告质粒的激活,甚至还有一些抑制作用。表达质粒pBFVORF-2对质粒pBFV-IP-4-luc没有激活。当补充pBOFR-1(1~249aa)时激活活性有所恢复(228倍),但也低于2的激活水平,甚至低于4的激活水平。这一结果表明:BFV读码框架编码的蛋白中,除orf-1编码的Tas蛋白对IP起始基因表达有强烈的激活作用外,其它的蛋白并不参与IP的功能。而且其中一些蛋白对Tas的反式激活功能还有抑制作用,这种抑制称为反式显性抑制作用(trans-dominant inhibitory effect)。这一结论与LTR也是一致的[7]。这可能是因为突变型蛋白或非调节蛋白的表达(通常在细胞中浓度较高)封阻了调节蛋白的特异细胞内因子[10]。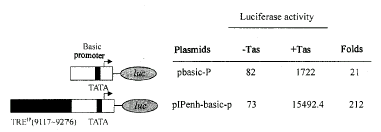
, http://www.100md.com
图2 BFV Tas对杂合启动子的激活作用
Figure 2 Transactivation of hybrid plasmid by BFV Tas
表2 BFV ORFs编码的蛋白对pBFV-IP-4-luc质粒的激活作用
Table 2 Transactivation of pBFV-IP-4-luc by protein encoded by BFV ORFs Cotransfected plasmida
Luc activity of
pBFV-IP-4-lucb
Folds
, 百拇医药
1 pUC19
108.5
-
2 pBORF-1(1~249aa)
31139.5
287
3 pBORF(1+2)
868
8
4 pBORF(1+2)+pBORF1(1~249aa)
26365.5
243
, http://www.100md.com
5 pBORF2
97.8
0.9
6 pBORF2+pBORF-1(1~249aa)
24738
228
7 pBORF1(1~217aa)
217
2
a.Background of plasmids were described in Materials and Methods 3.
, 百拇医药
b.Doses of plasmids were shown in Figure 1.
另外,从表2第7栏我们可以看出,C端缺失33aa的Tas[质粒pBORF1(1~217aa)]对于pBFV-IP-4-luc只有2倍的激活,说明Tas C端对于Tas激活IP的功能是至关重要的。
5 完整IP与LTR的同源性比较
以TATA盒为参照物,将完整IP与LTR部分区域进行比较,只发现极短簇的同源区。IP和LTR较低的同源性(27%)(图3),暗示了Tas对这两个启动子的反式激活机制可能不同,也许有不同的细胞因子参与。
图3 以TATA盒为参照物,完整IP与部分LTR序列的比较
, 百拇医药
Figure 3 Analytical comparison of intact IP with partial LTR sequence,using TATA box as reference Asterisk (*) indicates the identical nucleotide.
讨 论
我室前期工作表明BFV除了LTR外,在env基因3′端存在内部启动子IP[2]。本文证实IP启动子起始转录的功能受Tas调控。Tas调控位点包含于IP TATA盒上游160bp的序列之中(9?117~9?276)。本文已确定位于9?280~9?285之间的TATA盒为IP行使功能所必需。但Tas究竟是直接与IP结合还是通过与其它细胞因子间接作用于IP,尚需进一步的研究。研究表明:在SFV-1基因组8?171~8?176之间存在一个NF-1结合位点和糖皮质激素应答元件(GRE),但在HFV、FeFV中则没有[11]。BFV IP中也没有发现GRE元件,但在TREIP中存在细胞因子NF-1(nuclear factor 1)的结合位点(BFV 9?223~9?236)。这也说明了不同泡沫病毒的Tas调控机制可能是不同的,而且这种调控有可能是通过Tas与IP上特定序列结合而实现的。IP上的Tas应答元件仍需精确定位,其作用机制也有待进一步的研究。
, 百拇医药
BFV IP与LTR TATA盒附近序列只有一些散在的同源序列。暗示这两种元件是通过不同的机制被反式激活的。泡沫病毒反式激活因子Tas复杂的结构域为研究这一机制提供了基础。HFV、SFV、BFV的反式激活因子分别被称为Bel-1、Taf、Borf-1,它们与LTR调控相关功能域均已研究清楚,包括:(a)N-末端影响反式激活的效率;(b)中部含核定位信号和DNA结合域;(c)C-末端为反式激活所必需的激活域。从本文的结果看,Tas C-末端32aa对于Tas激活IP是必不可少的,但N-末端及其它结构域是否也对IP起同样的作用,以及作用的方式尚不清楚,需要通过进一步的缺失或定点突变来研究这种机制。
反式显性抑制作用在HIV-1、HTLV中均有报道[12,13],在HFV中不仅Bel-l的某些突变型具有这种作用,而且Bet、Bet突变型,甚至Bel-3、Beo等蛋白对Bel-l的反式激活功能都具有不同程度的显性抑制作用[14,15,16],这是因为Bel-l某些突变型或Bet等蛋白在细胞内过量表达以异源二聚体或混合多聚体的形式封阻了Bel-l入核所需的细胞因子;或者是这些多聚体有的能入核而占据了Bell所需靶点[15]。本研究中发现,Borf-1突变型(1~217aa)及pBFVORF(1+2)部分表达产物不同程度的抑制Borf-1对IP的反式激活作用,其机理可能与HFV一致。
, http://www.100md.com
基金项目:国家自然科学基金(39770032);天津市自然科学基金(983607511)
作者简介:张莉(1973-),女,湖南人,分子遗传学硕士。
联系作者:耿运琪,南开大学生命科学学院教授。
参考文献:
[1]刘佳建,刘淑红,陈启民,等.牛泡沫病毒编码的ORF-1为自身的反式激活因子[J].科学通报,1999,44(8):844-847.
[2]张莉,王世珍,刘佳建,等.牛泡沫病毒内部启动子(IP)的克隆及功能分析[J].病毒学报,2000,16(3):227-230.
[3]刘淑红,陈荷新,陈家童,等.牛泡沫病毒(BSV)3026毒株的分离及分子生物学鉴定[J].病毒学报,1997,13(2):140-145.
, http://www.100md.com
[4] Lochelt M, Flugel R M, Zboud M, et al. Human foamy virus genome possesses an internal, Bel-1-dependent and functional promoter [J] . Proc Natl Acad Sci USA, 1993, 90: 7317-7321.
[5] Mergia A. Simian foamy virus type 1 contains a second promoter located at the 3' end of the env gene [J] . Virology, 1994, 199: 219-222.
[6] Donald L, Holzschu, Mari A, et al. The nucleotide sequence and spliced pol mRNA levels of the nonprimate spumavirus bovine foamy virus [J] . J Virol, 1998, 72 (3) : 2177-2182.
, 百拇医药
[7]刘佳建.牛泡沫病毒(BFV)的基因表达调控及其与BIV的相互作用[D].天津:南开大学,1999.
[8] Sambrook J, Fritsch E F, Maniatis T. Molecular Cloning: A Laboratory Manual [M] , 2 ed, New York, USA: Cold Spring Harbor Laboratory Press, 1989.
[9]刘国文,纪永刚,梁臣,等.萤火虫荧光素酶基因构建BIV-LTR启动子表达体系[J].病毒学报,1994,10(4):377-380.
[10] Ptashne M, Gann A F. Activators and targets [J] . Nature, 1990, 346: 329-331.
[11] Campell M, Renshaw-gegg L, Renne R, et al. Characterization of the internal promoter of simian foamy viruses [J] . J Virol, 1994, 68: 4811-4820.
, http://www.100md.com
[12] Herskowits I. Functional inaction of genes by dominant negative mutations [J] . Nature, 1987, 329: 219-22.
[13] Baltimore D. Intracellular immunization [J] . Nature, 1990, 335: 395-396.
[14] He F, Sun J D, Garrett E D, et al. Functional organization of the bel-1 transactivator of human foamy virus [J] . J Virol, 1993, 67: 1896-1904.
[15] Venkatesh L K, Yang C, Theodorakis P A, et al. Functional dissection of the human spumaretrovirus transactivator identifies distinct classes of dominant-negative mutants [J] . J Virol, 1993, 67: 161-169.
[16] Baunach G, Maurer B, Hahn H, et al. Functional analysis of human foamy virus accessory reading frames [J] . J Virol, 1993, 67: 5411-5418.
收稿日期:1999-05-18;修回日期:1999-11-29, 百拇医药
单位:南开大学生命科学学院,天津 300071
关键词:牛泡沫病毒;内部启动子(IP);转录反式激活因子(Tas);反式激活因子应答元件(TRE);TATA盒
病毒学报000309 摘要:泡沫病毒的基因转录依赖至少两个不同的启动子:LTR调节病毒结构蛋白的表达,而内部启动子(IP)则起始调节蛋白mRNA的转录。牛泡沫病毒(BFV)在env与3′LTR之间有两个重叠的开放阅读框架orf-1和orf-2,分别编码BFV ORF-1、ORF-2等多种调节蛋白。这些蛋白中BFV ORF-1为转录激活因子,称为Tas。Tas对LTR及IP均有反式激活作用。BFV中第二类启动子IP的存在反映了泡沫病毒基因调控的复杂性。已从BFV3026中国毒株中克隆了基因组区段8 572~9 509,并通过测序和功能分析证明该区段包含了BFV IP。为了对IP进行更精确的定位,对其进行了进一步的缺失分析,将完整的IP定位在9 117~9 405之间。该启动子的TATA盒位于91 280~9 285,是IP必不可少的元件之一。杂合启动子和缺失分析表明,TATA盒上游160bp的区域内包含了IP的Tas应答元件(TREIP),其功能类似于增强子。Tas对IP具有强烈的激活作用,而C端缺失的ORF-1蛋白或ORF-2蛋白均不足以激活IP的表达。
, 百拇医药
中图分类号:S852.65+9.3; Q75 文献标识码:A
文章编号:1000-8721(2000)03-0232-06
Regulatory Elements That Mediate Transactivation by Tas in the Internal Promoter of Bovine Foamy Virus
ZHANG Li,QIAO Wen-tao,LIU Shu-hong,WANG Jin-zhon,CHEN Qi-min,GENG Yun-qi
(College of Life Sciences, Nankai University, Tianjin 300071, China)
Abstract: Foamy viruses are unusual among retroviruses in that they contain two distinct promoter elements. They are the long terminal repeat (LTR) promoter which regulates expression of the viral structural proteins, and a second internal promoter that locates towards the 3' end of the env gene and directs expression of the viral auxiliary proteins. One of these auxiliary proteins is a potent transcriptional transactivator, termed Tas in BFV. That is critical for foamy virus replication. Previously, we have cloned the intact BFV IP and demonstrated it to be highly responsive to the Tas protein. Here, we extended this earlier work by using deleted mutants and hybrid construct to localize the IP more precisely and to identify and characterize the Tas responsive element (TREIP) . Based on these data, we propose that genomic region 9117~9405nt contains all the sequences required for full Tas responsiveness. The importance of a TATA box for the BFV internal promoter is established by deletion analysis. The cis-acting sequences in the BFV env gene required for response to Tas are contained within a 160-bp element located 5' to the TATA box in the internal promoter. This Tas responsive element in the internal promoter functions in an orientation-independent fashion in a heterologous promoter and thus has the properties of an enhancer which depends on Tas activity. Alignments reveal that the BFV internal promoter and the BFV LTR have little sequence relatedness. In summary, the genome of bovine foamy virus directs viral transcription through both the promoter in the LTR and an internal promoter within the env gene, and each promoter contains unique enhancer-like elements regulated by the viral transactivator. And it seems that the Tas is the only accessory BFV protein necessary for viral replication in vitro.
, 百拇医药
Key words: bovine foamy virus; internal promoter; Tas; Tas responsive element; long terminal repeat promoter
泡沫病毒属于复杂的反转录病毒,它们不仅有复杂的基因组结构,而且具有复杂的基因表达调控机制。泡沫病毒自身编码一个病毒复制所必需的转录反式激活因子Tas。Tas可极大地激活病毒5′LTR起始的基因表达,而且这种激活作用在各种哺乳类乃至禽类细胞中均存在。BFV为非灵长类泡沫病毒,在env和3′LTR之间有两个开放阅读框架(ORF),其中第一个ORF编码BFV Tas蛋白[1]。Tas通过LTR U3区的多个顺式调控位点调节基因表达。本室以前的研究证明除了LTR外,在BFV env基因3′端存在另一个转录启动子,称为内部启动子[2],其活性亦强烈依赖于Tas。IP的存在使泡沫病毒基因表达调节机制比其它反转录病毒更显复杂。所以研究IP是研究泡沫病毒基因调控机制的一个重要环节。为了找到IP的精确位置,同时为了探寻BFV Tas在IP上的靶序列,本文构建了一系列IP缺失质粒以及IP/LTR杂合质粒,将它们转染牛肺细胞系BL-12,引入luc基因作瞬时表达分析。结果将完整的IP定位于基因组9?117~9?405之间。
, 百拇医药
材料与方法
1 病毒Hirt DNA的提取 反转录病毒感染宿主细胞后首先反转录成双链DNA,在反转录病毒感染的细胞的细胞核内常常存在一些非整合的线形或环状双链DNA分子,通常称为Hirt DNA。按文献[3]的方法从被BFV3026病毒株感染的新生牛肺细胞(NBL)中抽提NBL/BFV3026的Hirt DNA,并用TE缓冲液溶解。
2 引物的设计与PCR反应 参考有关HFV[4]和SFV-1[5]文献及最新发表的BFV基因组序列[6],根据env基因中的保守序列设计多对引物(表1),旨在扩增IP的缺失质粒。PCR反应条件为:94℃ 3min,50℃ 1min,72℃ 1min,1个循环;94℃ 30s,50℃ 1min,72℃ 1min,30个循环;94℃ 30s,50℃ 1min,72℃ 10min,1个循环。
, 百拇医药
3 缺失质粒的构建 质粒pBFV-IP-1-luc为本室构建[2],已证明该质粒上的8?572~9?509基因组片段包含了完整的牛泡沫病毒内部启动子。以此质粒为出发质粒,通过PCR方法构建了一系列IP缺失质粒pBFV-IP(2~6)-luc。质粒pBORF-1(1~249aa)、pBORF(1+2)、pBORF2、pBORF1(1~217aa)、pbasic-P由本室构建[7]。pBORF-1(1~249aa)表达完整的BFV ORF-1蛋白,即BFV的反式激活因子Tas;pBORF(1+2)表达orf-1、orf-2所编码的ORF-1、ORF-2、Bet等;pBORF-1(1~217aa)表达ORF-1蛋白的1~217aa肽段;pBFV ORF-2表达完整的BFV ORF-2蛋白(335aa)。这些表达质粒的上游启动子均为强启动子RSV LTR。pbasic-P是将BFV LTR的基本启动子(BFV基因组843~1012区段)克隆在luc基因上游[7]。 质粒pUC19、 pCMV-lacZ、 宿主菌E.coli JM103、TG1均由本室保存。
, 百拇医药
表1 构建质粒采用的引物
Table 1 Primers used in plasmid construction Plasmid
Actual genomic regions
contained in the plasmid
Upper/lower primers
pBFVIP-1-luc
BFV bp 8572~9509
P8564-8592:5′-ATA GGA AGT CGA CTG GAC TGG TCA GTA CT-3′
, http://www.100md.com SalⅠ
P9488-9517:5′-GAT AAG CTT TAT CCC AAT AAT GAC CGG TTT-3
HindⅢ
pBFVIP-2-luc
BFV bp 9053~9509
P9048-90655′ TCA TGT CGA CCC ACA CTG 3′
SalⅠ
p9488-9517:5′-GAT AAG CTT TAT CCC AAT AAT GAC CGG TTT-3′
HindⅢ
, 百拇医药
pBFVIP-3-luc
BFV bp 9053~9405
P9048-9065 5′ TCA TGT CGA C CC ACA CTG 3′
SalⅠ
P9395-9414 5′ CTC AAA GCT TAC GCA GCT AC 3′
HindⅢ
pBFVIP-4-luc
BFV bp 9117~9405
P9112-9130 5′ CTC TGT CGA CTG CTA TGGT 3′
, 百拇医药
SalⅠ
P9395-9414 5′ CTC AAA GCT TAC GCA GCT AC 3′
HindⅢ
pBFVIP-5-luc
BFV bp 9205~9405
P9200-9218 5′ AGA GGT CGA CTT CCT CATT 3′
SalⅠ
P9395-9414 5′ CTC AAA GCT TAC GCA GCT AC 3′
HindⅢ
pBFVIP-6-luc
, http://www.100md.com
BFV bp 9053~9276
P9048-9065 5′ TCA TGT CGA CCC ACA CTG 3′
SalⅠ
P9267-9285 5′ TTT AAA GCT TCC CAA GTGC 3′
HindⅢ
4 酶和试剂 实验用限制性内切酶、E.coli DNA聚合酶Ⅰ(K.F)、T4连接酶购自华美生物工程公司。荧光素酶检测试剂盒购自美国Promega公司。DMEM细胞培养基、青霉素/链霉素(p/s)、hepes及Trypsin购自美国GIBCO/BRL公司。
5 质粒的提取、酶切、DNA片段制备、连接及转化 其操作均参照文献[8]。
, 百拇医药
6 细胞培养和转染 用含10%胎牛血清的DMEM,37℃5%CO2培养。NBL细胞用于病毒繁殖,BL-12细胞系用于转染。转染时,BL-12以2.5×105ml-1接种于6孔板,每孔2ml。转染采用阳离子脂质体法(LipofectAMINE Reagent)。具体转染步骤参见GIBCO/BRL产品说明书。转染除用pUC19作对照质粒外,均设有β-gal内对照系统。ONPG、β-galactosidase购自Sigma公司。2天后裂解细胞测定Luc酶活力。细胞裂解液和Luc酶活力测定试剂盒购自Promega公司。按试剂盒提供的方法,用生物化学发光测量仪(上海检测技术所检测仪器厂生产)测定化学发光强度,方法原理见文献[9]。Luc酶的活力以4S内的光强度表示。结 果
1 完整的IP位于基因组9?117~9?405之间
本室以前的工作已证实了BFV env基因3′末端(基因组8?564~9?517区段)编码一个可被Tas激活的内部启动子[2]。PCR扩增的该区段被插入到无启动子的萤火虫荧光素酶基因上游构成表达质粒pBFV-IP-1-luc。Luc分析表明,单独转染时,pBFV-IP-1-luc只有很低的基础Luc活性,但当与pBORF-1共转染时,活性上升了145倍(图1),这说明IP的活性强烈依赖于Tas。
, http://www.100md.com
为了将IP在基因组上的位置定位得更加精细,我们以质粒pBFV-IP-1-luc为出发质粒,构建了IP的一系列缺失体pBFV-IP(2~6)-luc。将它们分别单独转染或与pBORF-1质粒共转染BL-12细胞系,转染结果如图1所示。克隆片段的相对大小及它们在基因组中的位置在图中亦有表示。由于这些缺失体的基础活性近似,所以推断IP中不含负调控区。当下游保持在9?509,上游缺失481 bp至基因组9?053时(pBFV-IP-2-luc),激活倍数不但没有降低,反而大大提高,由此可以推断缺失掉的这481 bp不影响Tas对IP的激活作用,是无关序列;而当上游为9?053,下游缺失至9?405时(pBFV-IP-3-luc),激活程度并不低于Tas对pBFV-IP-2-luc的作用,所以同样说明下游9?405~9?509这段序列也与IP的功能关系不大。我们此时对上游进行进一步的缺失至9?117(pBFV-IP-4-luc),发现激活倍数达到了350倍。而若继续缺失至9?205时(pBFV-IP-5-luc),激活活性则有所下降。说明IP的上限应定在9?117。在这一基础上,我们对IP的下游又进行了缺失,直至9?276。从图1可看出,激活倍数降到了最低水平3.6,与其它质粒相比可认为没有激活。由此可见,下游9?276~9?405也是IP行使功能必不可少的。因此,我们将IP的下限定在9?405。综上所述,我们可将完整的IP定位于BFV基因组9?117~9?405之间。
, 百拇医药
图1 IP系列缺失体及转染BL-12细胞系的Luc分析结果
Figure 1 A series of IP deleted mutants and the outcome of Luc assay when transfected them into BL-12 cell line
A.Schematic diagram of BFV IP and the amplificated genome regions by PCR;luc.firefly luciferase gene(upper).Lines represent BFV IP regions remained after deletion.B.“-Tas”means cotransfection of the reporter plasmid and pUC19.This value represents basal promoter activity.“+Tas”means cotransfection of the reporter plasmid and the transactivator plasmid (pBFVORF-1).a.Each experiment was done at least 3 times in triplicate and each plate was transfected with 0.2μg of deleted luciferase expression plasmid, 0.75μg of pBFVORF-1,0.6μg of pCMV β-gal and 3 μg of Lipofect AMINE(GIBCO/BRL).The cells were harvested to determine Luc activity at 48h after transfection.Luciferase activities were normalized for β-galactosidase.Generally,less than 15% variation in replicate samples was observed.
, http://www.100md.com
2 Tas对IP的激活作用必须有TATA盒的参与
序列分析和同源比较表明:与BFV IP行使功能有重要关系的9?276~9?405这段区域中含有典型的真核启动子TATA盒。与其它泡沫病毒相比,该TATA盒及其附近的序列十分保守,均为TATAAAAGA*CA*AT。而除TATA盒外,9?276~9?405中未发现其它元件。所以IP的TATA盒是起始转录不可缺少的元件。这个结论同时也证实了IP起始转录是通过RNA pol Ⅱ复合体与TATA盒作用而实现的。
3 BFV IP上的Tas应答元件功能类似于增强子
为了深入研究Borf-1激活IP起始基因表达调控的特点,并且为了探寻Tas在IP上的作用位点,本文对IP TATA盒上游160bp的序列(9?117~9?276)与Borf-1之间的作用特点作了进一步的研究。质粒pbasic-P在luc基因上游含有BFV LTR的基本启动子,但缺少增强子元件,所以这个载体可用于研究增强子[7]。利用此质粒,我们构建了IP-basic P杂合启动子。BFV IP TATA盒近上游160bp的序列被克隆到LTR基本启动子(basic promoter)上游构建杂合体pIPenh-basic-P。瞬时分析实验表明,单独转染杂合体时Luc活性不高。但当它与激活质粒pBORF-1共转染时,Luc活性上升了212倍(图2),而且其激活活性也远远高于pbasic-P与pBORF-1共转染时活性。由此可见:IP TATA盒上游160bp序列强烈依赖于Tas,且有可能以增强子的方式发挥作用。结合杂合启动子以及图1中的缺失分析,我们确认这160bp IP序列中含有Tas作用元件TREIP。
, 百拇医药
4 Borf-1蛋白是IP起始基因表达唯一所需的病毒辅助蛋白
本室以前的工作已证明,BFV LTR起始转录依赖于BFV反式激活因子Tas蛋白,而ORF-2蛋白或C端缺失的ORF-1蛋白均不足以激活BFV LTR的表达[7]。从表2可以看出,这个结论同样适用于IP。本文采用野生型IP质粒pBFV-IP-4-luc作为报告质粒,分别与pBORF-1(1~249aa)、pBORF(1+2)、pBORF2、pBORF1(1~217aa)共转染BL-12,结果见表2。将pBFV-IP-4-luc与无关质粒pUC19共转染时的Luc值设为负对照。当将表达完整Tas的pBORF-1(1~249aa)质粒与pBFV-IP-4-luc共转染时,激活倍数为287;而当pBORF(1+2)与报告质粒共转染时,激活倍数只有8。即使在体系中再加入pBORF-1(1~249aa),激活活性也恢复不到pBORF1(1~249aa)单独与pBFV-IP-4-luc共转染时的水平。说明pBFVORF(1+2)表达产物中有部分蛋白不参与对报告质粒的激活,甚至还有一些抑制作用。表达质粒pBFVORF-2对质粒pBFV-IP-4-luc没有激活。当补充pBOFR-1(1~249aa)时激活活性有所恢复(228倍),但也低于2的激活水平,甚至低于4的激活水平。这一结果表明:BFV读码框架编码的蛋白中,除orf-1编码的Tas蛋白对IP起始基因表达有强烈的激活作用外,其它的蛋白并不参与IP的功能。而且其中一些蛋白对Tas的反式激活功能还有抑制作用,这种抑制称为反式显性抑制作用(trans-dominant inhibitory effect)。这一结论与LTR也是一致的[7]。这可能是因为突变型蛋白或非调节蛋白的表达(通常在细胞中浓度较高)封阻了调节蛋白的特异细胞内因子[10]。
, http://www.100md.com
图2 BFV Tas对杂合启动子的激活作用
Figure 2 Transactivation of hybrid plasmid by BFV Tas
表2 BFV ORFs编码的蛋白对pBFV-IP-4-luc质粒的激活作用
Table 2 Transactivation of pBFV-IP-4-luc by protein encoded by BFV ORFs Cotransfected plasmida
Luc activity of
pBFV-IP-4-lucb
Folds
, 百拇医药
1 pUC19
108.5
-
2 pBORF-1(1~249aa)
31139.5
287
3 pBORF(1+2)
868
8
4 pBORF(1+2)+pBORF1(1~249aa)
26365.5
243
, http://www.100md.com
5 pBORF2
97.8
0.9
6 pBORF2+pBORF-1(1~249aa)
24738
228
7 pBORF1(1~217aa)
217
2
a.Background of plasmids were described in Materials and Methods 3.
, 百拇医药
b.Doses of plasmids were shown in Figure 1.
另外,从表2第7栏我们可以看出,C端缺失33aa的Tas[质粒pBORF1(1~217aa)]对于pBFV-IP-4-luc只有2倍的激活,说明Tas C端对于Tas激活IP的功能是至关重要的。
5 完整IP与LTR的同源性比较
以TATA盒为参照物,将完整IP与LTR部分区域进行比较,只发现极短簇的同源区。IP和LTR较低的同源性(27%)(图3),暗示了Tas对这两个启动子的反式激活机制可能不同,也许有不同的细胞因子参与。
图3 以TATA盒为参照物,完整IP与部分LTR序列的比较
, 百拇医药
Figure 3 Analytical comparison of intact IP with partial LTR sequence,using TATA box as reference Asterisk (*) indicates the identical nucleotide.
讨 论
我室前期工作表明BFV除了LTR外,在env基因3′端存在内部启动子IP[2]。本文证实IP启动子起始转录的功能受Tas调控。Tas调控位点包含于IP TATA盒上游160bp的序列之中(9?117~9?276)。本文已确定位于9?280~9?285之间的TATA盒为IP行使功能所必需。但Tas究竟是直接与IP结合还是通过与其它细胞因子间接作用于IP,尚需进一步的研究。研究表明:在SFV-1基因组8?171~8?176之间存在一个NF-1结合位点和糖皮质激素应答元件(GRE),但在HFV、FeFV中则没有[11]。BFV IP中也没有发现GRE元件,但在TREIP中存在细胞因子NF-1(nuclear factor 1)的结合位点(BFV 9?223~9?236)。这也说明了不同泡沫病毒的Tas调控机制可能是不同的,而且这种调控有可能是通过Tas与IP上特定序列结合而实现的。IP上的Tas应答元件仍需精确定位,其作用机制也有待进一步的研究。
, 百拇医药
BFV IP与LTR TATA盒附近序列只有一些散在的同源序列。暗示这两种元件是通过不同的机制被反式激活的。泡沫病毒反式激活因子Tas复杂的结构域为研究这一机制提供了基础。HFV、SFV、BFV的反式激活因子分别被称为Bel-1、Taf、Borf-1,它们与LTR调控相关功能域均已研究清楚,包括:(a)N-末端影响反式激活的效率;(b)中部含核定位信号和DNA结合域;(c)C-末端为反式激活所必需的激活域。从本文的结果看,Tas C-末端32aa对于Tas激活IP是必不可少的,但N-末端及其它结构域是否也对IP起同样的作用,以及作用的方式尚不清楚,需要通过进一步的缺失或定点突变来研究这种机制。
反式显性抑制作用在HIV-1、HTLV中均有报道[12,13],在HFV中不仅Bel-l的某些突变型具有这种作用,而且Bet、Bet突变型,甚至Bel-3、Beo等蛋白对Bel-l的反式激活功能都具有不同程度的显性抑制作用[14,15,16],这是因为Bel-l某些突变型或Bet等蛋白在细胞内过量表达以异源二聚体或混合多聚体的形式封阻了Bel-l入核所需的细胞因子;或者是这些多聚体有的能入核而占据了Bell所需靶点[15]。本研究中发现,Borf-1突变型(1~217aa)及pBFVORF(1+2)部分表达产物不同程度的抑制Borf-1对IP的反式激活作用,其机理可能与HFV一致。
, http://www.100md.com
基金项目:国家自然科学基金(39770032);天津市自然科学基金(983607511)
作者简介:张莉(1973-),女,湖南人,分子遗传学硕士。
联系作者:耿运琪,南开大学生命科学学院教授。
参考文献:
[1]刘佳建,刘淑红,陈启民,等.牛泡沫病毒编码的ORF-1为自身的反式激活因子[J].科学通报,1999,44(8):844-847.
[2]张莉,王世珍,刘佳建,等.牛泡沫病毒内部启动子(IP)的克隆及功能分析[J].病毒学报,2000,16(3):227-230.
[3]刘淑红,陈荷新,陈家童,等.牛泡沫病毒(BSV)3026毒株的分离及分子生物学鉴定[J].病毒学报,1997,13(2):140-145.
, http://www.100md.com
[4] Lochelt M, Flugel R M, Zboud M, et al. Human foamy virus genome possesses an internal, Bel-1-dependent and functional promoter [J] . Proc Natl Acad Sci USA, 1993, 90: 7317-7321.
[5] Mergia A. Simian foamy virus type 1 contains a second promoter located at the 3' end of the env gene [J] . Virology, 1994, 199: 219-222.
[6] Donald L, Holzschu, Mari A, et al. The nucleotide sequence and spliced pol mRNA levels of the nonprimate spumavirus bovine foamy virus [J] . J Virol, 1998, 72 (3) : 2177-2182.
, 百拇医药
[7]刘佳建.牛泡沫病毒(BFV)的基因表达调控及其与BIV的相互作用[D].天津:南开大学,1999.
[8] Sambrook J, Fritsch E F, Maniatis T. Molecular Cloning: A Laboratory Manual [M] , 2 ed, New York, USA: Cold Spring Harbor Laboratory Press, 1989.
[9]刘国文,纪永刚,梁臣,等.萤火虫荧光素酶基因构建BIV-LTR启动子表达体系[J].病毒学报,1994,10(4):377-380.
[10] Ptashne M, Gann A F. Activators and targets [J] . Nature, 1990, 346: 329-331.
[11] Campell M, Renshaw-gegg L, Renne R, et al. Characterization of the internal promoter of simian foamy viruses [J] . J Virol, 1994, 68: 4811-4820.
, http://www.100md.com
[12] Herskowits I. Functional inaction of genes by dominant negative mutations [J] . Nature, 1987, 329: 219-22.
[13] Baltimore D. Intracellular immunization [J] . Nature, 1990, 335: 395-396.
[14] He F, Sun J D, Garrett E D, et al. Functional organization of the bel-1 transactivator of human foamy virus [J] . J Virol, 1993, 67: 1896-1904.
[15] Venkatesh L K, Yang C, Theodorakis P A, et al. Functional dissection of the human spumaretrovirus transactivator identifies distinct classes of dominant-negative mutants [J] . J Virol, 1993, 67: 161-169.
[16] Baunach G, Maurer B, Hahn H, et al. Functional analysis of human foamy virus accessory reading frames [J] . J Virol, 1993, 67: 5411-5418.
收稿日期:1999-05-18;修回日期:1999-11-29, 百拇医药