牛免疫缺陷病毒反式激活因子功能域的研究
作者:荣丽玮 王世珍 刘淑红 陈启民 耿运琪 C. Wood
单位:荣丽玮 王世珍 刘淑红 陈启民 耿运琪(南开大学生命科学院,天津 300071);C. Wood(美国Nebraska大学生命科学院,美国)
关键词:牛免疫缺陷病毒; 反式激活因子; 缺失突变
中国病毒学000115 分类号:Q75 文献识别码:A
文章编号:1003-5125(2000)01-0088-05
Study of Bovine Immunodeficiency Virus Transactivator
Functional Domains
, 百拇医药
RONG LI-wei,WANG Shi-zhen,LIU Shu-hong,CHEN Qi-min,GENG Yun-qi,Charles Wood
(College of Life Science, Nankai University, Tianjin 300071, China)
Abstract:Transactivator (Tat) of bovine immunodeficiency virus (BIV) is a virus-encoded regulatory protein, which activates gene expression directed by BIV long terminal repeat (LTR), and plays an important role in BIV replica tive cycle. With methods of fusion protein expression and deletion mutation, as et of BIV Tat deletion mutants was constructed, and co-transfected into FBL cells with BIV LTR. Using luciferase gene as a reporter, the function of BIV Tat mutants was detected and it showed that: 17/96 aa region of BIV Tat peptide contains full activity; auxiliary amino acids exist in N-terminal 17/34 aa region; Cys-rich region and basi c region are essential to BIV Tat activity. Computer prediction of BIV Tat secon dary structure was showed, and mechanism how it functions was analyzed.
, 百拇医药
Key words:Bovine immunodeficiency virus (BIV); Transactivator (Tat); Deletion mutation▲
牛免疫缺陷病毒(Bovine immunodeficiency virus, BIV)与人免疫缺陷病毒(Human immunod eficiency virus, HIV)同属反转录病毒科慢病毒属[1]。BIV基因组5′端的长末 端重复序列(LTR)起始病毒结构基因和非结构基因的转录[2],因而许多细胞因子 和病毒编码的调节蛋白作用于LTR,以调节BIV的基因表达。其中Tat蛋白是BIV的反式激活因 子,可大大提高LTR的转录水平,在BIV的基因表达及基因组复制的调节中起重要作用 [3]。HIV、马传染性贫血病毒(Equine infectious anemia virus, EIAV)和猴免疫 缺陷病毒(Simian immunodeficiency virus, SIV)的Tat蛋白均含有相同或相似的保守结构 功能域,即Cys富含区(Cys-rich region)、核心区(core region)和碱性区(basic region) [4~6]。氨基酸序列同源性比较表明,BIV Tat蛋白也具有上述三个保守功能域 [7],但目前尚缺乏BIV Tat蛋白功能域的直接实验证据。本研究通过缺失突变的 方法研究了BIV Tat蛋白不同功能域的作用,并通过模拟蛋白的二级结构,对各功能域的作 用机制进行了分析。
, 百拇医药
1 材料与方法
1.1 质粒构建 利用17aa和96aa附近的两个Hin dⅢ 位点,从克隆有完整BIV Tat(103个氨基酸)的pB-Tat(美国Nebraska大学Dr. Wood赠送)得 到pTat1(含BIV Tat蛋白17/96肽段)。此后分别利用34aa附近的Sty Ⅰ位点和45aa附近的Nde Ⅰ位点,由pTat1出发,得到pTat2(含BIV Tat蛋白34/96肽段)和pTat3(含BIV Tat蛋白45/96 肽段)。将pTat1、pTat2、pTat3中的Tat基因分别克隆至pD-181-luc[含有BIV LTR(-181 +204),其下游连有报告基因luc]中取代luc基因XbaⅠ位点下游的部分,并维持正确的读码 框 以表达融合Tat蛋白,得到pL-Tat1(表达BIV Tat 17/96肽段融合蛋白)、pL-Tat2(表达BIV Tat 34/96肽段融合蛋白)和pL-Tat3(表达BIV Tat 45/96肽段融合蛋白)。利用78aa附近的 BamHI位点由pL-Tat1构建得到pL-Tat4(表达BIV Tat 17/78肽段融合蛋白)。
, 百拇医药
1.2 细胞培养和转染 小牛肺细胞(Fetal bovine lung cells, FBL),用补加10%胎牛血清(购自天津生化制品厂)的DMEM培养基(购自GIBCO/BRL)于 37℃、5%CO2条件下培养。采用脂质体介导法转染细胞(参见GIBCO/BRL产品说明书)。
1.3 荧光素酶活性测定 转染开始后48h裂解细 胞,测定荧光素酶活性(参见Promega公司荧光素酶检测试剂盒操作说明)。
2 结果与讨论
2.1 系列BIV Tat缺失突变型对BIV LTR的激活作用
质粒pL-Tat1、pL-Tat2、pL-Tat3、pL-Tat4所编码的BIV Tat各缺失突变型如图1所示。 将各缺失质粒与pBIVLTR-luc共转染FBL细胞。pBIVLTR-luc克隆有含BIV Tat蛋白靶序列的 完整的BIV LTR启动子,及在该启动子控制下的荧光素酶基因。通过检测缺失前后转染细胞 裂解物荧光素酶活性的变化,可得知各BIV Tat缺失突变型激活功能的变化。以pBIVLTR -lu c和pUC18共转染FBL为本底,以pBIVLTR-luc和pB-Tat共转染FBL所得的luc激活倍数为参照 ,检测各BIV Tat缺失突变型产生的激活倍数的变化,结果如表1。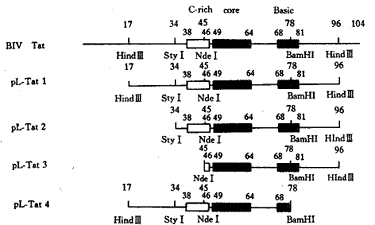
, 百拇医药
图1 BIV Tat蛋白序列缺失图谱 (图 中标出了所用的限制性酶切位点)
Fig.1 BIV Tat protein partial deletion map (Restriction enzymes sites are showe d).
表1 各BIV Tat缺失突变型对BIV LTR的激活作用
Table 1 Activation of BIV LTR by BIV Tat deletion mutants 转染用质粒a
Plasmids in transfection
荧光素酶活性b
Luciferase activity
, 百拇医药
激活倍数
Activation fold
-
6.7±0.6
-
pBIVLTR-luc+pUC18
62.7±13.8
-
pBIVLTR-luc+pB-Tat
1381±290
22.0
pBIVLTR-luc+pL-Tat1
, 百拇医药
1320±174
21.1
pBIVLTR-luc+pL-Tat2
408±54
6.5
pBIVLTR-luc+pL-Tat3
68.3±18.0
1.1
pBIVLTR-luc+pL-Tat4
59.7±14.6
95
a每种质粒DNA用量200ng,以脂质体法转染FBL细胞。b每组转染重复3次,转 染开始后48小时裂解细胞测定荧光素酶活性,计算3次的平均值。
, 百拇医药
aFBL cells were transfected by lipofect method with plasmids DNA in an amo unt of 200ng. bEach transfection was repeated 3 times. Cells were lysed and luciferase activity was examined 48 hours after transfection, and averages w ere calculated.
与全长103个氨基酸的野生型BIV Tat相比,pL-Tat1保留了17/96肽段,激活pBIVLTR-luc 的程度(21.1倍)与野生型(22.0倍)相似,说明17/96肽段拥有野生型Tat的全部激活功能,可 被初步界定为BIV Tat蛋白的功能肽段。当进一步缺失17/34肽段后(即pL-Tat2),34/96肽 段虽然仍含有完整的三个功能域,但激活倍数由野生型Tat的22.0倍下降至6.5倍,说明保守 功能域以外的17/34肽段对于BIV Tat的激活功能也有一定作用,可能在这一区域中有些辅助 氨基酸起到稳定RNA-蛋白质复合体的功能。在pL-Tat3表达的45/96肽段中,Cys富含区所 含的5个保守半胱氨酸缺失了4个,该肽段不再具有激活功能,显示了Cys富含区的重要作 用。在BIV Tat中,该区段很可能与在HIV-1 Tat中一样,直接参与激活作用。在pL-Tat4 表达的17/78肽段中,碱性区因Arg80和Arg81缺失而遭到破坏,导致激活功 能完全丧失。这两个精氨酸很可能参与了Tat蛋白与靶序列的结合。进一步的研究将确定更 为精细的BIV Tat功能图谱及核心区的作用。
, 百拇医药
对HIV-1 Tat的研究已经证实,Cys富含区中半胱氨酸的突变导致Tat功能的完全丧失;核心 区和Cys富含区一起,对于HIV-1 Tat行使激活功能是必不可少的。BIV Tat的这两个功能结 构域与HIV-1的同源性为54%,其中核心区序列的同源性高达70%。BIV Tat的碱性区与HIV- 1也具有一定的同源性,HIV中该区域是使Tat定位于细胞核的元件,并通过一个精氨酸与靶 序列的环区特异性识别。由于BIV Tat与HIV-1 Tat的结构功能极为相似,对其进行深入研 究将有助于阐明慢病毒基因表达调控机制。
2.2 Tat蛋白的二级结构模拟
计算机模拟蛋白的二级结构如图2所示:肽链N端有一段α螺旋(5/11aa),但它对蛋白活性不 是必需的。17/34肽段中含有一段β折叠,并有4个芳香族氨基酸,推测它们的侧链可嵌入RN A碱基之间,在蛋白与其靶序列结合时稳定RNA-蛋白质复合物。Cys富含区(38/46aa)与后面 的核心区(49/63aa)共同构成肽链中央一段较长的β折叠,前者使Tat形成有活性的二聚体形 式,并可能与核心区一起参与激活作用,或者与某些二价金属离子结合,共同构成激活核心 区。碱性区(68/81aa)则表现为β转角的构象,与其周围的几个氨基酸形成一个三个转角组 合的“弓”字,它决定了蛋白与RNA的结合。碱性氨基酸带有正电荷,可提供与RNA结合的表 面。BIV LTR中Tat的靶序列RNA形成独有的茎环结构,富含精氨酸的碱性区采取β转角的空 间构象可嵌合于靶序列RNA的大沟中,该区域的核磁共振波谱学研究也证明了这一假说 [8]。我们猜测Tat蛋白可沿RNA双螺旋滑动,直至遇到其靶序列上的一个凸出部位,蛋 白和靶序列同时发生某种构象变化,碱性区内的氨基酸完成对该凸出部位的识别,蛋白由非 特异性结合转为序列特异性结合。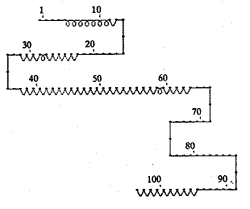
, http://www.100md.com
图2 BIV Tat二级结构预测示意图
Fig.2 Schematic diagram of BIV Tat secondary structure prediction
对Tat蛋白带电荷情况和疏水性进行分析发现:该蛋白存在两个富含正电荷的区域,位于Cys 富 含区上游的32/40肽段(含有4个正电荷)和碱性区(含有8个正电荷);它们之间夹着一段疏水 性的核心区。这样的结构暗示着两边带正电的区域可与靶序列表面结合,并进行特异性的识 别;蛋白被锚定后,中间的疏水区伸进RNA双螺旋结构的内部,引起RNA解链或其他二级结构 变化,并可能由其他蛋白因子介导,实现其激活功能。■
基金项目:国家自然科学基金(39670032)和美国NIH资助国际合作项目(R03 TW00493-01A1)
作者简介:荣丽玮(1971-),女,满族,籍贯陕西省,1996年毕业于南开大学生物化学与分子生物学系,分子遗传学硕士,现在加拿大McGill大学攻读实验医学博士学位。
, 百拇医药
耿运琪 (1947-),男,籍贯河北省,南开大学生命科学学院分子病毒学教授。
参考文献:
[1]Van Der Maaten M J, Boothe A D, Seger C L. Isolation of a virus f rom cattle with persistent lymphocytosis [J]. J Natl Cancer Inst, 1972,49:1649~1657
[2]Garvey K J, Oberste M S, Elser J E et al. Nucleotide sequence and genome organization of biologically active proviruses of the bovine immunodeficiency-like virus [J]. Virology, 1990,175:391~409
, 百拇医药
[3]Liu Z Q, Sheridan D, Wood C. Identification and characterization of the bovine immunodeficiency-like virus tat gene [J]. J Virol, 1992,66:5137~5140
[4]Ruben S, Perkins A, Purcell R et al. Structural and functional chara cterization of human immunodeficiency virus Tat protein [J]. J Virol, 1989, 63 :1~8
[5]Garcia J A. Functional domains required for Tat-induced transcriptional activation of the HIV-1 long terminal repeat. EMBO [J], 1988,6:3761~3770
, 百拇医药
[6]Cullen B R. Mechanism of action of regulatory proteins encoded by complex retroviruses [J]. Microbiol Rev, 1992,56:375~394
[7]荣丽玮.牛免疫缺陷病毒反式激活因子功能结构域的初步研究[D].[学位论文].天津:南开大学,1996.
[8]Puglisi J D, Chen L, Blanchard S et al. Solution structure of a bovine immunodeficiency virus Tat-TAR peptide-RNA complex [J]. Science,1995,270: 1200~1203
收稿日期:1998-11-23
修稿日期:1999-01-22, 百拇医药
单位:荣丽玮 王世珍 刘淑红 陈启民 耿运琪(南开大学生命科学院,天津 300071);C. Wood(美国Nebraska大学生命科学院,美国)
关键词:牛免疫缺陷病毒; 反式激活因子; 缺失突变
中国病毒学000115 分类号:Q75 文献识别码:A
文章编号:1003-5125(2000)01-0088-05
Study of Bovine Immunodeficiency Virus Transactivator
Functional Domains
, 百拇医药
RONG LI-wei,WANG Shi-zhen,LIU Shu-hong,CHEN Qi-min,GENG Yun-qi,Charles Wood
(College of Life Science, Nankai University, Tianjin 300071, China)
Abstract:Transactivator (Tat) of bovine immunodeficiency virus (BIV) is a virus-encoded regulatory protein, which activates gene expression directed by BIV long terminal repeat (LTR), and plays an important role in BIV replica tive cycle. With methods of fusion protein expression and deletion mutation, as et of BIV Tat deletion mutants was constructed, and co-transfected into FBL cells with BIV LTR. Using luciferase gene as a reporter, the function of BIV Tat mutants was detected and it showed that: 17/96 aa region of BIV Tat peptide contains full activity; auxiliary amino acids exist in N-terminal 17/34 aa region; Cys-rich region and basi c region are essential to BIV Tat activity. Computer prediction of BIV Tat secon dary structure was showed, and mechanism how it functions was analyzed.
, 百拇医药
Key words:Bovine immunodeficiency virus (BIV); Transactivator (Tat); Deletion mutation▲
牛免疫缺陷病毒(Bovine immunodeficiency virus, BIV)与人免疫缺陷病毒(Human immunod eficiency virus, HIV)同属反转录病毒科慢病毒属[1]。BIV基因组5′端的长末 端重复序列(LTR)起始病毒结构基因和非结构基因的转录[2],因而许多细胞因子 和病毒编码的调节蛋白作用于LTR,以调节BIV的基因表达。其中Tat蛋白是BIV的反式激活因 子,可大大提高LTR的转录水平,在BIV的基因表达及基因组复制的调节中起重要作用 [3]。HIV、马传染性贫血病毒(Equine infectious anemia virus, EIAV)和猴免疫 缺陷病毒(Simian immunodeficiency virus, SIV)的Tat蛋白均含有相同或相似的保守结构 功能域,即Cys富含区(Cys-rich region)、核心区(core region)和碱性区(basic region) [4~6]。氨基酸序列同源性比较表明,BIV Tat蛋白也具有上述三个保守功能域 [7],但目前尚缺乏BIV Tat蛋白功能域的直接实验证据。本研究通过缺失突变的 方法研究了BIV Tat蛋白不同功能域的作用,并通过模拟蛋白的二级结构,对各功能域的作 用机制进行了分析。
, 百拇医药
1 材料与方法
1.1 质粒构建 利用17aa和96aa附近的两个Hin dⅢ 位点,从克隆有完整BIV Tat(103个氨基酸)的pB-Tat(美国Nebraska大学Dr. Wood赠送)得 到pTat1(含BIV Tat蛋白17/96肽段)。此后分别利用34aa附近的Sty Ⅰ位点和45aa附近的Nde Ⅰ位点,由pTat1出发,得到pTat2(含BIV Tat蛋白34/96肽段)和pTat3(含BIV Tat蛋白45/96 肽段)。将pTat1、pTat2、pTat3中的Tat基因分别克隆至pD-181-luc[含有BIV LTR(-181 +204),其下游连有报告基因luc]中取代luc基因XbaⅠ位点下游的部分,并维持正确的读码 框 以表达融合Tat蛋白,得到pL-Tat1(表达BIV Tat 17/96肽段融合蛋白)、pL-Tat2(表达BIV Tat 34/96肽段融合蛋白)和pL-Tat3(表达BIV Tat 45/96肽段融合蛋白)。利用78aa附近的 BamHI位点由pL-Tat1构建得到pL-Tat4(表达BIV Tat 17/78肽段融合蛋白)。
, 百拇医药
1.2 细胞培养和转染 小牛肺细胞(Fetal bovine lung cells, FBL),用补加10%胎牛血清(购自天津生化制品厂)的DMEM培养基(购自GIBCO/BRL)于 37℃、5%CO2条件下培养。采用脂质体介导法转染细胞(参见GIBCO/BRL产品说明书)。
1.3 荧光素酶活性测定 转染开始后48h裂解细 胞,测定荧光素酶活性(参见Promega公司荧光素酶检测试剂盒操作说明)。
2 结果与讨论
2.1 系列BIV Tat缺失突变型对BIV LTR的激活作用
质粒pL-Tat1、pL-Tat2、pL-Tat3、pL-Tat4所编码的BIV Tat各缺失突变型如图1所示。 将各缺失质粒与pBIVLTR-luc共转染FBL细胞。pBIVLTR-luc克隆有含BIV Tat蛋白靶序列的 完整的BIV LTR启动子,及在该启动子控制下的荧光素酶基因。通过检测缺失前后转染细胞 裂解物荧光素酶活性的变化,可得知各BIV Tat缺失突变型激活功能的变化。以pBIVLTR -lu c和pUC18共转染FBL为本底,以pBIVLTR-luc和pB-Tat共转染FBL所得的luc激活倍数为参照 ,检测各BIV Tat缺失突变型产生的激活倍数的变化,结果如表1。
, 百拇医药
图1 BIV Tat蛋白序列缺失图谱 (图 中标出了所用的限制性酶切位点)
Fig.1 BIV Tat protein partial deletion map (Restriction enzymes sites are showe d).
表1 各BIV Tat缺失突变型对BIV LTR的激活作用
Table 1 Activation of BIV LTR by BIV Tat deletion mutants 转染用质粒a
Plasmids in transfection
荧光素酶活性b
Luciferase activity
, 百拇医药
激活倍数
Activation fold
-
6.7±0.6
-
pBIVLTR-luc+pUC18
62.7±13.8
-
pBIVLTR-luc+pB-Tat
1381±290
22.0
pBIVLTR-luc+pL-Tat1
, 百拇医药
1320±174
21.1
pBIVLTR-luc+pL-Tat2
408±54
6.5
pBIVLTR-luc+pL-Tat3
68.3±18.0
1.1
pBIVLTR-luc+pL-Tat4
59.7±14.6
95
a每种质粒DNA用量200ng,以脂质体法转染FBL细胞。b每组转染重复3次,转 染开始后48小时裂解细胞测定荧光素酶活性,计算3次的平均值。
, 百拇医药
aFBL cells were transfected by lipofect method with plasmids DNA in an amo unt of 200ng. bEach transfection was repeated 3 times. Cells were lysed and luciferase activity was examined 48 hours after transfection, and averages w ere calculated.
与全长103个氨基酸的野生型BIV Tat相比,pL-Tat1保留了17/96肽段,激活pBIVLTR-luc 的程度(21.1倍)与野生型(22.0倍)相似,说明17/96肽段拥有野生型Tat的全部激活功能,可 被初步界定为BIV Tat蛋白的功能肽段。当进一步缺失17/34肽段后(即pL-Tat2),34/96肽 段虽然仍含有完整的三个功能域,但激活倍数由野生型Tat的22.0倍下降至6.5倍,说明保守 功能域以外的17/34肽段对于BIV Tat的激活功能也有一定作用,可能在这一区域中有些辅助 氨基酸起到稳定RNA-蛋白质复合体的功能。在pL-Tat3表达的45/96肽段中,Cys富含区所 含的5个保守半胱氨酸缺失了4个,该肽段不再具有激活功能,显示了Cys富含区的重要作 用。在BIV Tat中,该区段很可能与在HIV-1 Tat中一样,直接参与激活作用。在pL-Tat4 表达的17/78肽段中,碱性区因Arg80和Arg81缺失而遭到破坏,导致激活功 能完全丧失。这两个精氨酸很可能参与了Tat蛋白与靶序列的结合。进一步的研究将确定更 为精细的BIV Tat功能图谱及核心区的作用。
, 百拇医药
对HIV-1 Tat的研究已经证实,Cys富含区中半胱氨酸的突变导致Tat功能的完全丧失;核心 区和Cys富含区一起,对于HIV-1 Tat行使激活功能是必不可少的。BIV Tat的这两个功能结 构域与HIV-1的同源性为54%,其中核心区序列的同源性高达70%。BIV Tat的碱性区与HIV- 1也具有一定的同源性,HIV中该区域是使Tat定位于细胞核的元件,并通过一个精氨酸与靶 序列的环区特异性识别。由于BIV Tat与HIV-1 Tat的结构功能极为相似,对其进行深入研 究将有助于阐明慢病毒基因表达调控机制。
2.2 Tat蛋白的二级结构模拟
计算机模拟蛋白的二级结构如图2所示:肽链N端有一段α螺旋(5/11aa),但它对蛋白活性不 是必需的。17/34肽段中含有一段β折叠,并有4个芳香族氨基酸,推测它们的侧链可嵌入RN A碱基之间,在蛋白与其靶序列结合时稳定RNA-蛋白质复合物。Cys富含区(38/46aa)与后面 的核心区(49/63aa)共同构成肽链中央一段较长的β折叠,前者使Tat形成有活性的二聚体形 式,并可能与核心区一起参与激活作用,或者与某些二价金属离子结合,共同构成激活核心 区。碱性区(68/81aa)则表现为β转角的构象,与其周围的几个氨基酸形成一个三个转角组 合的“弓”字,它决定了蛋白与RNA的结合。碱性氨基酸带有正电荷,可提供与RNA结合的表 面。BIV LTR中Tat的靶序列RNA形成独有的茎环结构,富含精氨酸的碱性区采取β转角的空 间构象可嵌合于靶序列RNA的大沟中,该区域的核磁共振波谱学研究也证明了这一假说 [8]。我们猜测Tat蛋白可沿RNA双螺旋滑动,直至遇到其靶序列上的一个凸出部位,蛋 白和靶序列同时发生某种构象变化,碱性区内的氨基酸完成对该凸出部位的识别,蛋白由非 特异性结合转为序列特异性结合。
, http://www.100md.com
图2 BIV Tat二级结构预测示意图
Fig.2 Schematic diagram of BIV Tat secondary structure prediction
对Tat蛋白带电荷情况和疏水性进行分析发现:该蛋白存在两个富含正电荷的区域,位于Cys 富 含区上游的32/40肽段(含有4个正电荷)和碱性区(含有8个正电荷);它们之间夹着一段疏水 性的核心区。这样的结构暗示着两边带正电的区域可与靶序列表面结合,并进行特异性的识 别;蛋白被锚定后,中间的疏水区伸进RNA双螺旋结构的内部,引起RNA解链或其他二级结构 变化,并可能由其他蛋白因子介导,实现其激活功能。■
基金项目:国家自然科学基金(39670032)和美国NIH资助国际合作项目(R03 TW00493-01A1)
作者简介:荣丽玮(1971-),女,满族,籍贯陕西省,1996年毕业于南开大学生物化学与分子生物学系,分子遗传学硕士,现在加拿大McGill大学攻读实验医学博士学位。
, 百拇医药
耿运琪 (1947-),男,籍贯河北省,南开大学生命科学学院分子病毒学教授。
参考文献:
[1]Van Der Maaten M J, Boothe A D, Seger C L. Isolation of a virus f rom cattle with persistent lymphocytosis [J]. J Natl Cancer Inst, 1972,49:1649~1657
[2]Garvey K J, Oberste M S, Elser J E et al. Nucleotide sequence and genome organization of biologically active proviruses of the bovine immunodeficiency-like virus [J]. Virology, 1990,175:391~409
, 百拇医药
[3]Liu Z Q, Sheridan D, Wood C. Identification and characterization of the bovine immunodeficiency-like virus tat gene [J]. J Virol, 1992,66:5137~5140
[4]Ruben S, Perkins A, Purcell R et al. Structural and functional chara cterization of human immunodeficiency virus Tat protein [J]. J Virol, 1989, 63 :1~8
[5]Garcia J A. Functional domains required for Tat-induced transcriptional activation of the HIV-1 long terminal repeat. EMBO [J], 1988,6:3761~3770
, 百拇医药
[6]Cullen B R. Mechanism of action of regulatory proteins encoded by complex retroviruses [J]. Microbiol Rev, 1992,56:375~394
[7]荣丽玮.牛免疫缺陷病毒反式激活因子功能结构域的初步研究[D].[学位论文].天津:南开大学,1996.
[8]Puglisi J D, Chen L, Blanchard S et al. Solution structure of a bovine immunodeficiency virus Tat-TAR peptide-RNA complex [J]. Science,1995,270: 1200~1203
收稿日期:1998-11-23
修稿日期:1999-01-22, 百拇医药