实验性帕金森病纹状体、大脑皮质、海马组织Glu含量的变化
作者:徐仁伵 孙圣刚 刘煜敏 童萼塘 梅元武
单位:徐仁伵(同济医科大学附属协和医院神经科 430022);孙圣刚(同济医科大学附属协和医院神经科 430022);童萼塘(同济医科大学附属协和医院神经科 430022);梅元武(同济医科大学附属协和医院神经科 430022);刘煜敏(湖北医科大学附属第二医院神经科)
关键词:帕金森病;谷氨酸
脑与神经疾病杂志000201 摘 要 目的:探索帕金森病(PD) 的发病机制。方法: 应用大鼠选择性偏侧(右侧)PD动物模型, 观 测正常对照组, PD组(造模后2周)左右侧纹状体(ST)、大脑皮质(CC)、海马(HP)组织谷氨酸 (Glu)含量的变化。 结果: PD组右侧ST、CC、HP组织Glu含量显著 低于左侧和正常对照组左右侧(P<0.01); PD组左侧ST、CC、HP组织Glu含 量与正常对照组比 较无显著差异性(P>0.05); 正常对照组左右侧ST、CC、HP组织Glu含量无显著 差异性(P>0.0 5)。 结论: 神经递质Glu参于PD累及部位ST、CC、HP损伤的发病机 制
, http://www.100md.com
The changes of striatum,cerebral cortex and hippocampus Glu content on experimental Parkinson's disease
Xu Renshi,Sun Shenggang,Liu Yimei,et al
Department of Neurology,Union Hosital of Tongji Medical University.
Abstract Objective: To investigate the mechanism of Parkinson's disease (PD). Methods: The contents of striatum (ST), cerebral cortex (CC) and hippocampus (HP) gluta mate (Glu) was examined on rat control group, raight lateral PD model group (2 w eeks). Results:The right lateral contents of ST, CC and HP Glu on rat PD gro up decrease significantly than that of the PD group left and the control group b o th side (P<0.01). The PD group left ST, CC and HP Glu contents don't obviously c hange, compared with that of the control's (P>0.05). The control gro up two later al ST, CC and HP Glu contents hasn't significant difference (P>0.05) . C onclusion: Neurotransmitter Glu acts on the mechanism of PD striatum, c erebral cortex, hippocampus damage.
, 百拇医药
Key words Parkinson's disease(PD) Glutamate(Glu)
帕金森病(Parkinson's disease,PD)是一种以中脑黑质多巴胺(DA)能神经元严重缺失为 主要病变, 广泛累及中枢神经系的较常见的、 症候复 杂多样的慢性神经元变性性疾病。 目前就其发病机制尚无定论, 本课题想通过观测PD累及 部位纹状体(Striatum, ST)、 大脑皮质(Cerebral cortex, CC)、 海马(Hippocampus, HP) 组织谷氨酸(Glutamate, Glu)含量的变化, 试着探索PD发病的可能机制。
材 料 和 方 法
一、 选择性偏侧大鼠PD模型制备
1. 手术前准备: 成熟健康wistar大白鼠, 雌雄不拘, 体重150~200g。 动物在室温下 笼中常规喂养, 术前不禁食。 6-羟多巴胺(6-OHDA.HBr, Sigma chemial Co.产品)溶于 0 .02%的抗坏血酸液中配成0.2%的溶液, 低温(-20℃)冰箱密封避光保存备用。 阿朴吗啡( Ap omorphine.HCl,APO, Sigma chemical Co.产品)用双蒸水配成0.01%溶液备用, 以上溶 液要求严格无菌。
, 百拇医药
2. 选择性右侧偏侧大鼠PD模型制作[1,2]; 动物用0.3%戊巴比妥钠10ml/kg腹 腔注射麻醉后, 固定在立体定位仪上(江湾Ⅰ型C)。 在无菌条件下, 正中切开大鼠颅顶部皮肤, 剥离骨膜, 暴露前囱(十字缝)、 人字缝。 用鼠颅骨钻于手术要求部位钻一直径为 5mm的颅骨孔。 参照Pellegrino等[3]大鼠脑立体定位图谱, 参看Whishaw等(图谱 附有)体重在150-200g范围内的鼠使用此图谱的数学折算方法。 确定右侧黑质部坐标位置 。 用10μl微量进样器(上海计量仪器厂生产)分两次将6-OHDA按下列坐标位置注入右侧黑 质部建立右侧选择性大鼠PD动物模型。 B坐标系统: 外耳道连线(耳杆尖中心连线)低于上 门齿板上缘5mm, 即水平0平面。 第一次注射4μl, 针尖斜面朝嘴侧, 坐标为, 前囱后(P os terior bregma, PB)3.0mm, 正中线右侧(Median right lateral, MRL)2.5mm, 硬脑膜 腹侧 (Ventraldura, VD)8.6mm。 第二次注射3μl, 针尖斜面朝尾侧, 坐标为, PB2.4mm, MR L2.7mm, VD8.6mm。 注射速度1μl/min, 每次注射完毕留针10min, 然后以1.0mm/min速 度缓慢拔针(约10min)。 手术完毕, 用医用明胶海绵填塞颅骨孔, 缝合切口, 放回笼中 喂养。
, 百拇医药
3. APO诱发动物旋转行为观察: 造模术后2周, 动物腹腔注射0.01%APO10ml/kg诱发大鼠 旋转行为(逆时针方向), 验证6-OHDA对中脑黑质DA能神经元的损伤效应。 旋转速度达到6 r/min(35cm/r)视为成功PD选择性偏侧模型, 注射APO 10min开始记录, 共记录30min。
二、 动物分组及处理 实验动物分为正常对照组和PD模型组, 每组8只。 PD组选择符合 实验要求的于第2周迅速断头处死, 快速取脑, 在冰盘中分离双侧ST、 CC、 HP组织, 称 湿重。 测定Glu的标本用4℃双蒸水匀浆, 碱性条件下用2.4-二硝基氟苯高效液相色谱仪 柱前衍生色谱法测定, 色谱条件: 流动相A为CH3CN: H2O(1∶1), 流动相B为0.05ml /L的NaAc-HAc缓冲液, 流速0.7ml/min, 柱温30℃, 检测波长UV-360nm。 Glu标准品购 自 Sigma chemical Co., LC-6A高效液相色谱仪, SPD-6AV紫外分光光度计均为日本岛津公 司产品。 正常对照组除不行手术外, 其余处理方法与PD组相同。
, 百拇医药
三、 统计学处理 数据以均数±标准差表示“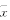 ±s”。 组间比较采用t 检验, 检验水准a=0.05。
±s”。 组间比较采用t 检验, 检验水准a=0.05。
结 果
PD组右侧ST、CC和HP组织Glu含量显著低于左侧和正常对照组左右侧(P<0.01); PD 组左侧ST、CC和HP组Glu含量与正常对照组左侧比较无显著差异性(P>0.05); 正常对照组左右侧ST、CC和HP组织Glu含量无显著差异性(P>0.05), 见表1。
表1 正常对照组、PD组鼠ST、CC和HP组织Glu含量(m g/g Wt)
正常对照组
PD组
左侧
, 百拇医药
右侧
左侧
右侧
ST组织Glu
5.60±1.58
5.58±1.49△
5.97±1.66△
3.42±1.20*
CC组织Glu
3.39±0.78
3.41±1.50△
, 百拇医药
3.40±0.93△
1.25±0.33*
HP组织Glu
7.51±1.05
7.40±1.90△
7.56±1.17△
5.44±0.6 5*
n=8 *与PD组左侧、正常对照组左右比较,P<0.01
△与正常对照组左侧比较,P>0.05
讨 论
, 百拇医药
一、 脑组织Glu的含量与Glu能神经元Glu递质释放的关系。
Glu有代谢库与递质库 之分,代谢库是指存在于胞浆及线粒体中的部分, 递质库是指贮存于末梢突触小泡内的Glu , 造成细胞外液Glu浓度持续升高的原因有: 摄取功能下降, 甚至摄取方向的逆转及末梢 释放增加。 Glu的释放受多种因素的调节, 除离子依赖性, 电压依赖性通道的开闭等调节 外, 还受神经递质的调节, 如DA、 GABA、 腺苷、 甘氨酸等。 组织Glu释放增加, 大部 分聚积在突触间隙或弥散到细胞外液, 细胞外液中多数在此聚积, 少数经血液淋巴液丢失 。 所以在组织Glu代谢库和递质库不变的情况下, 神经元Glu释放越多, 则组织Glu下降越 明显, 反之也然[4~6].
二、 Glu对神经元的损伤作用。
Glu是中枢神经系统中兴奋性神经元突触的主要神经递质, 同时又是一种神经毒素。 Glu 大量释放和/或重摄取障碍, Glu在细胞外液中堆聚产生兴奋毒性作用。 Glu造成的神经元 兴奋毒性作用主要有两个过程: 一是以Na+、 Cl-、 H2O的内流, 使细胞水肿为特 征的急性损伤过程; 二是通过激活Glu受体, 直接或间接启动Ca2+通道, Ca2+ 大量内流, Na+-Ca2+交换减弱, 以及IP3使细胞贮存Ca2+释放增加, 细胞内外Ca2+平衡紊乱, 细胞内Ca2+超载造成迟发性神经元变性、 坏死的迟 发性损伤过程。 在大多数病理情况下, 尤其是中枢性神经系统慢性进行性疾病, 如PD等 , NMDA受体过度兴奋导致的神经元迟发性变性、 坏死可能与PD发病具有密切联系[ 7,8], 其间有氧自由基代谢异常参与介导。 Ca2+还可通过激活NMDA受体产生一 氧化氮直接杀伤神经元[9]。
, http://www.100md.com
三、 PD发病机制的探讨。
锥体外系结构十分复杂, 其纤维联系相当广泛, 除与运动调控有关的ST、 CC、 丘脑、 黑质神经信息反馈回路外, 还与小脑齿状核, 中脑红核, 桥脑脑桥核、 前庭核、 端脑 海马结构, 脊髓前角细胞等具有直接或间接的纤维联系。 这些神经元间的纤维联系牵涉许 多递质: 确知者已有多巴胺、 乙酰胆碱、 γ-氨基丁酸、 Glu、 5-羟色胺、 去甲肾上 腺素、 P物质、 阿片肽等。 不同性能的神经元通过这些递质(或调质)相互拮抗、 调整、 以维持功能的平衡。 在PD中, 递质的平衡遭受破坏, 直接引发症候或间接引发和递质密 切相关的突触后受体、 神经元受损间接引发PD症候。 研究表明[10,11], Glu的 大多数递质通路与基底核(ST)、 CC、 边缘系统(海马结构)有关, 其兴奋毒性作用与PD的 发生机理密切相关。 实验中我们观测到: PD组动物黑质损伤侧(右侧)ST、CC和HP组织Glu 含量显著低于黑质未损伤侧(左侧)和正常对照组左、 右侧; PD组左侧ST、 CC和HP组织Glu 与正常对照组比较无显著差异; 正常对照组左、 右侧ST、CC和HP组织Glu含量无显著差异 。 我们推测: 神经递质Glu参与PD累及部位ST、CC和HP损伤的发病机制。 其机制可能是: 黑质DA能神经元严重变性、 坏死, ST、CC和HP直接或间接失神经支配, 神经递质平衡失 调(代偿性的升高或降低), Glu释放增加, 引起ST、CC和HP组织Glu能神经元受损引发其功 能相关的PD症候。
, 百拇医药
本课题系国家教委统招博士研究生课题基金资助
参考文献
1 Perese DA, Ulman J, Viola J, et al. A 6-hydroxydopa mine-induced selective parkinsonian rat model. Brain Res, 1989;494∶285.
2 Thomas J, Wang J, Takubo H, et al. A 6-hydroxydopamine-induced se lective parkinsonian rat model: further biochemical and behavioral characterizat on. Exp Neurol, 1994;126∶159.
3 Pellegrino LJ, Pellegrino AS, Cushman AJ. A stereotaxic atlas of the rat brain. 2nd ed. Plenum press, 1979;43.
, 百拇医药
4 Desce JM, Godeheu G, Galli T, et al. Presynaptic facilitation of dopa mine release through D, L-alpha-amino-3-hydroxy-5-methyl-4-isoxazole pro pionate receptors on synaptosomes from the rat striatum. J Pharmacol Exp Ther, 1991;259∶692.
5 Hortnagl H, Berger ML, Reither H, et al. Cholinergic deficit induced by ethylcholine aziridinum (AF64A) in rat hippocampus: effect of glutamatergic s ystems. Arch Pharmacol, 1991;344∶213.
6 Gannon RL, Terrian DM. Presynaptic modulation of glutamate and dynorp hin release by excitatory amino acids in the guinea-pig hippocampus. Neuroscien ce, 1991;41∶401.
, http://www.100md.com
7 Whetsell WO. Current concept of excitotoxicity. Neurology, 1996;55(1) ∶1.
8 Kikuchi, Kim SU. Glutamate neurotoxicity on mesencephalic dopaminergi c neurons in culture. J Neurosci Res, 1993;36∶558.
9 岳少杰, 虞佩兰. 谷氨酸的兴奋性神经毒性作用与神经系统损伤. 国外医学神 经病学神经外科学分册, 1994;21(4)∶205.
10 Porras A, Mora F. Dopamine-glutamate-GABA interaction and agine: studies in the striatum of the conscious rat. Eur J Neurosci, 1995;7∶2183.
11 Kavler R, Calder LD, Thai LH, et al. The dopaminergic, glutamatergic , GABAergic bases for the action of amphetamine and cocaine. Brain Res, 1995;671 ∶100.
(1999-11-15 收稿), http://www.100md.com
单位:徐仁伵(同济医科大学附属协和医院神经科 430022);孙圣刚(同济医科大学附属协和医院神经科 430022);童萼塘(同济医科大学附属协和医院神经科 430022);梅元武(同济医科大学附属协和医院神经科 430022);刘煜敏(湖北医科大学附属第二医院神经科)
关键词:帕金森病;谷氨酸
脑与神经疾病杂志000201 摘 要 目的:探索帕金森病(PD) 的发病机制。方法: 应用大鼠选择性偏侧(右侧)PD动物模型, 观 测正常对照组, PD组(造模后2周)左右侧纹状体(ST)、大脑皮质(CC)、海马(HP)组织谷氨酸 (Glu)含量的变化。 结果: PD组右侧ST、CC、HP组织Glu含量显著 低于左侧和正常对照组左右侧(P<0.01); PD组左侧ST、CC、HP组织Glu含 量与正常对照组比 较无显著差异性(P>0.05); 正常对照组左右侧ST、CC、HP组织Glu含量无显著 差异性(P>0.0 5)。 结论: 神经递质Glu参于PD累及部位ST、CC、HP损伤的发病机 制
, http://www.100md.com
The changes of striatum,cerebral cortex and hippocampus Glu content on experimental Parkinson's disease
Xu Renshi,Sun Shenggang,Liu Yimei,et al
Department of Neurology,Union Hosital of Tongji Medical University.
Abstract Objective: To investigate the mechanism of Parkinson's disease (PD). Methods: The contents of striatum (ST), cerebral cortex (CC) and hippocampus (HP) gluta mate (Glu) was examined on rat control group, raight lateral PD model group (2 w eeks). Results:The right lateral contents of ST, CC and HP Glu on rat PD gro up decrease significantly than that of the PD group left and the control group b o th side (P<0.01). The PD group left ST, CC and HP Glu contents don't obviously c hange, compared with that of the control's (P>0.05). The control gro up two later al ST, CC and HP Glu contents hasn't significant difference (P>0.05) . C onclusion: Neurotransmitter Glu acts on the mechanism of PD striatum, c erebral cortex, hippocampus damage.
, 百拇医药
Key words Parkinson's disease(PD) Glutamate(Glu)
帕金森病(Parkinson's disease,PD)是一种以中脑黑质多巴胺(DA)能神经元严重缺失为 主要病变, 广泛累及中枢神经系的较常见的、 症候复 杂多样的慢性神经元变性性疾病。 目前就其发病机制尚无定论, 本课题想通过观测PD累及 部位纹状体(Striatum, ST)、 大脑皮质(Cerebral cortex, CC)、 海马(Hippocampus, HP) 组织谷氨酸(Glutamate, Glu)含量的变化, 试着探索PD发病的可能机制。
材 料 和 方 法
一、 选择性偏侧大鼠PD模型制备
1. 手术前准备: 成熟健康wistar大白鼠, 雌雄不拘, 体重150~200g。 动物在室温下 笼中常规喂养, 术前不禁食。 6-羟多巴胺(6-OHDA.HBr, Sigma chemial Co.产品)溶于 0 .02%的抗坏血酸液中配成0.2%的溶液, 低温(-20℃)冰箱密封避光保存备用。 阿朴吗啡( Ap omorphine.HCl,APO, Sigma chemical Co.产品)用双蒸水配成0.01%溶液备用, 以上溶 液要求严格无菌。
, 百拇医药
2. 选择性右侧偏侧大鼠PD模型制作[1,2]; 动物用0.3%戊巴比妥钠10ml/kg腹 腔注射麻醉后, 固定在立体定位仪上(江湾Ⅰ型C)。 在无菌条件下, 正中切开大鼠颅顶部皮肤, 剥离骨膜, 暴露前囱(十字缝)、 人字缝。 用鼠颅骨钻于手术要求部位钻一直径为 5mm的颅骨孔。 参照Pellegrino等[3]大鼠脑立体定位图谱, 参看Whishaw等(图谱 附有)体重在150-200g范围内的鼠使用此图谱的数学折算方法。 确定右侧黑质部坐标位置 。 用10μl微量进样器(上海计量仪器厂生产)分两次将6-OHDA按下列坐标位置注入右侧黑 质部建立右侧选择性大鼠PD动物模型。 B坐标系统: 外耳道连线(耳杆尖中心连线)低于上 门齿板上缘5mm, 即水平0平面。 第一次注射4μl, 针尖斜面朝嘴侧, 坐标为, 前囱后(P os terior bregma, PB)3.0mm, 正中线右侧(Median right lateral, MRL)2.5mm, 硬脑膜 腹侧 (Ventraldura, VD)8.6mm。 第二次注射3μl, 针尖斜面朝尾侧, 坐标为, PB2.4mm, MR L2.7mm, VD8.6mm。 注射速度1μl/min, 每次注射完毕留针10min, 然后以1.0mm/min速 度缓慢拔针(约10min)。 手术完毕, 用医用明胶海绵填塞颅骨孔, 缝合切口, 放回笼中 喂养。
, 百拇医药
3. APO诱发动物旋转行为观察: 造模术后2周, 动物腹腔注射0.01%APO10ml/kg诱发大鼠 旋转行为(逆时针方向), 验证6-OHDA对中脑黑质DA能神经元的损伤效应。 旋转速度达到6 r/min(35cm/r)视为成功PD选择性偏侧模型, 注射APO 10min开始记录, 共记录30min。
二、 动物分组及处理 实验动物分为正常对照组和PD模型组, 每组8只。 PD组选择符合 实验要求的于第2周迅速断头处死, 快速取脑, 在冰盘中分离双侧ST、 CC、 HP组织, 称 湿重。 测定Glu的标本用4℃双蒸水匀浆, 碱性条件下用2.4-二硝基氟苯高效液相色谱仪 柱前衍生色谱法测定, 色谱条件: 流动相A为CH3CN: H2O(1∶1), 流动相B为0.05ml /L的NaAc-HAc缓冲液, 流速0.7ml/min, 柱温30℃, 检测波长UV-360nm。 Glu标准品购 自 Sigma chemical Co., LC-6A高效液相色谱仪, SPD-6AV紫外分光光度计均为日本岛津公 司产品。 正常对照组除不行手术外, 其余处理方法与PD组相同。
, 百拇医药
三、 统计学处理 数据以均数±标准差表示“
±s”。 组间比较采用t 检验, 检验水准a=0.05。结 果
PD组右侧ST、CC和HP组织Glu含量显著低于左侧和正常对照组左右侧(P<0.01); PD 组左侧ST、CC和HP组Glu含量与正常对照组左侧比较无显著差异性(P>0.05); 正常对照组左右侧ST、CC和HP组织Glu含量无显著差异性(P>0.05), 见表1。
表1 正常对照组、PD组鼠ST、CC和HP组织Glu含量(m g/g Wt)
正常对照组
PD组
左侧
, 百拇医药
右侧
左侧
右侧
ST组织Glu
5.60±1.58
5.58±1.49△
5.97±1.66△
3.42±1.20*
CC组织Glu
3.39±0.78
3.41±1.50△
, 百拇医药
3.40±0.93△
1.25±0.33*
HP组织Glu
7.51±1.05
7.40±1.90△
7.56±1.17△
5.44±0.6 5*
n=8 *与PD组左侧、正常对照组左右比较,P<0.01
△与正常对照组左侧比较,P>0.05
讨 论
, 百拇医药
一、 脑组织Glu的含量与Glu能神经元Glu递质释放的关系。
Glu有代谢库与递质库 之分,代谢库是指存在于胞浆及线粒体中的部分, 递质库是指贮存于末梢突触小泡内的Glu , 造成细胞外液Glu浓度持续升高的原因有: 摄取功能下降, 甚至摄取方向的逆转及末梢 释放增加。 Glu的释放受多种因素的调节, 除离子依赖性, 电压依赖性通道的开闭等调节 外, 还受神经递质的调节, 如DA、 GABA、 腺苷、 甘氨酸等。 组织Glu释放增加, 大部 分聚积在突触间隙或弥散到细胞外液, 细胞外液中多数在此聚积, 少数经血液淋巴液丢失 。 所以在组织Glu代谢库和递质库不变的情况下, 神经元Glu释放越多, 则组织Glu下降越 明显, 反之也然[4~6].
二、 Glu对神经元的损伤作用。
Glu是中枢神经系统中兴奋性神经元突触的主要神经递质, 同时又是一种神经毒素。 Glu 大量释放和/或重摄取障碍, Glu在细胞外液中堆聚产生兴奋毒性作用。 Glu造成的神经元 兴奋毒性作用主要有两个过程: 一是以Na+、 Cl-、 H2O的内流, 使细胞水肿为特 征的急性损伤过程; 二是通过激活Glu受体, 直接或间接启动Ca2+通道, Ca2+ 大量内流, Na+-Ca2+交换减弱, 以及IP3使细胞贮存Ca2+释放增加, 细胞内外Ca2+平衡紊乱, 细胞内Ca2+超载造成迟发性神经元变性、 坏死的迟 发性损伤过程。 在大多数病理情况下, 尤其是中枢性神经系统慢性进行性疾病, 如PD等 , NMDA受体过度兴奋导致的神经元迟发性变性、 坏死可能与PD发病具有密切联系[ 7,8], 其间有氧自由基代谢异常参与介导。 Ca2+还可通过激活NMDA受体产生一 氧化氮直接杀伤神经元[9]。
, http://www.100md.com
三、 PD发病机制的探讨。
锥体外系结构十分复杂, 其纤维联系相当广泛, 除与运动调控有关的ST、 CC、 丘脑、 黑质神经信息反馈回路外, 还与小脑齿状核, 中脑红核, 桥脑脑桥核、 前庭核、 端脑 海马结构, 脊髓前角细胞等具有直接或间接的纤维联系。 这些神经元间的纤维联系牵涉许 多递质: 确知者已有多巴胺、 乙酰胆碱、 γ-氨基丁酸、 Glu、 5-羟色胺、 去甲肾上 腺素、 P物质、 阿片肽等。 不同性能的神经元通过这些递质(或调质)相互拮抗、 调整、 以维持功能的平衡。 在PD中, 递质的平衡遭受破坏, 直接引发症候或间接引发和递质密 切相关的突触后受体、 神经元受损间接引发PD症候。 研究表明[10,11], Glu的 大多数递质通路与基底核(ST)、 CC、 边缘系统(海马结构)有关, 其兴奋毒性作用与PD的 发生机理密切相关。 实验中我们观测到: PD组动物黑质损伤侧(右侧)ST、CC和HP组织Glu 含量显著低于黑质未损伤侧(左侧)和正常对照组左、 右侧; PD组左侧ST、 CC和HP组织Glu 与正常对照组比较无显著差异; 正常对照组左、 右侧ST、CC和HP组织Glu含量无显著差异 。 我们推测: 神经递质Glu参与PD累及部位ST、CC和HP损伤的发病机制。 其机制可能是: 黑质DA能神经元严重变性、 坏死, ST、CC和HP直接或间接失神经支配, 神经递质平衡失 调(代偿性的升高或降低), Glu释放增加, 引起ST、CC和HP组织Glu能神经元受损引发其功 能相关的PD症候。
, 百拇医药
本课题系国家教委统招博士研究生课题基金资助
参考文献
1 Perese DA, Ulman J, Viola J, et al. A 6-hydroxydopa mine-induced selective parkinsonian rat model. Brain Res, 1989;494∶285.
2 Thomas J, Wang J, Takubo H, et al. A 6-hydroxydopamine-induced se lective parkinsonian rat model: further biochemical and behavioral characterizat on. Exp Neurol, 1994;126∶159.
3 Pellegrino LJ, Pellegrino AS, Cushman AJ. A stereotaxic atlas of the rat brain. 2nd ed. Plenum press, 1979;43.
, 百拇医药
4 Desce JM, Godeheu G, Galli T, et al. Presynaptic facilitation of dopa mine release through D, L-alpha-amino-3-hydroxy-5-methyl-4-isoxazole pro pionate receptors on synaptosomes from the rat striatum. J Pharmacol Exp Ther, 1991;259∶692.
5 Hortnagl H, Berger ML, Reither H, et al. Cholinergic deficit induced by ethylcholine aziridinum (AF64A) in rat hippocampus: effect of glutamatergic s ystems. Arch Pharmacol, 1991;344∶213.
6 Gannon RL, Terrian DM. Presynaptic modulation of glutamate and dynorp hin release by excitatory amino acids in the guinea-pig hippocampus. Neuroscien ce, 1991;41∶401.
, http://www.100md.com
7 Whetsell WO. Current concept of excitotoxicity. Neurology, 1996;55(1) ∶1.
8 Kikuchi, Kim SU. Glutamate neurotoxicity on mesencephalic dopaminergi c neurons in culture. J Neurosci Res, 1993;36∶558.
9 岳少杰, 虞佩兰. 谷氨酸的兴奋性神经毒性作用与神经系统损伤. 国外医学神 经病学神经外科学分册, 1994;21(4)∶205.
10 Porras A, Mora F. Dopamine-glutamate-GABA interaction and agine: studies in the striatum of the conscious rat. Eur J Neurosci, 1995;7∶2183.
11 Kavler R, Calder LD, Thai LH, et al. The dopaminergic, glutamatergic , GABAergic bases for the action of amphetamine and cocaine. Brain Res, 1995;671 ∶100.
(1999-11-15 收稿), http://www.100md.com