长春花叶片愈伤组织诱导及培养过程中生物碱合成类型的变化和调节及有关酶的分析
作者:赵 剑* 朱蔚华 胡 秋 王文科
单位:赵 剑* 朱蔚华 胡 秋 中国协和医科大学药物研究所 北京 100050中国医学科学院;王文科 山西师范大学生物系
关键词:长春花愈伤组织;吲哚生物碱;过氧化物酶
中草药990729 摘 要 在 4 种不同培养基上诱导长春花叶片愈伤组织发生的过程中,叶片中含量较高的文多灵(vindoline)和长春质碱(catharanthine)迅速降解至极低水平,而在叶片中含量较低的阿玛碱(ajmalicine)和蛇根碱(serpentine)的合成却逐渐增加,同时酸性和碱性过氧化物酶(peroxidase)活性呈上升趋势,尤其碱性过氧化物酶与蛇根碱和阿玛碱的相关性较密切。光照和黑暗中的变化趋势基本一致,但光照可提高过氧化物酶的活性,对培养物的长春质碱、文多灵和蛇根碱的合成有利。不同的培养基对生物碱的合成影响较大,2,4-D 显著抑制生物碱的合成,持续光照使愈伤组织的生长受到抑制,使阿玛碱向其蛇根碱转化。
, 百拇医药
Regulation of Indole Alkaloids Synthesis and Related Enzymes During Callus
Initiation and Culture of Madagascar Periwinkle Leaves (Catharanthus roseus)
Zhao Jian, Wang Wenke, Zhu Weihua, et al
(Institute of Materia Medica of PUMC & CAMC, Beijing 100050)
Abstract Leaf calluses of Catharanthus roseus (L.) G.Don were initiated in four different media under illumination or in dark. Visible callus began to form within 10 to 15 days. The ordinarily high contents of vindoline and catharanthine in the leaf declined rapidly, while the lower ajmalicine and serpentine contents increased markedly during callus initiation. At the same time, acidic and alkaline peroxidase showed a tendency to increase their activities, especially the activity of alkaline peroxidase was more closely related with the synthesis of serpentine and ajmalicine. Results also showed that illumination improved occurrence of leaf callus in advance and stimulated peroxidase activity, beneficial for the synthesis of vindoline, catharanthine and serpentine. 2.4-D represses biosynthesis of indole alkaloids and peroxidase activity in all cultures.
, 百拇医药
Key words Catharanthus roseus (L.) G.Don leaf callus initiation indole alkaloid peroxidase activity
长春花组织和细胞培养已经研究了很多年,但至今仍仅限于中等规模的生物反应器生产,且长春花生物碱的生物合成途径和调节有许多不清楚之处,所以国内外的很多实验室仍在进行研究[1~3]。尤其近几年在长春碱(vinblastine)的基础上得到新的半合成抗癌药异长春碱(vinorelbine)具有很大的应用潜力,市场价格高于紫杉醇。而其半合成原料仍需长春质碱和文多灵,使组织培养这一可能的原料来源途径再次受到人们关注。从长春花愈伤组织的诱导、继代培养到细胞悬浮培养这个过程中培养物的生物碱合成代谢类型和能力有了很大变化,这一切是如何发生的还很不清楚,本文试图从愈伤组织的诱导和继代培养过程中的生物碱合成类型的变化和调节及此过程中的相关酶活性变化来探索其原因。
, http://www.100md.com
1 实验材料和方法
1.1 愈伤组织的诱导:取实验室内盆栽的长春花顶芽下第三片叶,用肥皂水和自来水清洗干净,吸干水分,先用70 %酒精灭菌2 min 后,转入0.1 %的升汞灭菌10 min,无菌蒸馏水冲洗5次,剪成小块,接种于以下4种诱导培养基中:M1:MS+NAA 1 mg/L+IAA 1 mg/L+KT 0.1 mg/L+5%蔗糖+0.7%琼脂;M2:MS+NAA 1 mg/L+KT 0.1 mg/L+5%蔗糖+0.7%琼脂;M3:MS+2,4-D 1 mg/L+KT 0.1 mg/L+3%蔗糖+0.7%琼脂;L1:LS+IAA 1 mg/L+6-BA 0.1 mg/L+5%蔗糖+0.7%琼脂,培养基在高温灭菌前均调至pH8.5。每瓶接种物约为0.4 g。分别置于光下和黑暗中培养,培养温度均为 23℃±2℃,光照周期12 h/d,光强约2 000 lx。激素和光的调节实验用来自同一植株的组织系,取继代3次的该愈伤组织均匀地接种于含有不同种类和浓度的生长素和激动素组合的MS培养基含3 %蔗糖和0.7 %琼脂(激素配比编号为 A,B,C,D,E,F),分别置于光下和黑暗中培养,培养基制备和培养条件与以上相同。愈伤组织诱导实验每隔5 d取一次材料,光和激素调节实验则在培养45 d后取材,称鲜重后,一部分做酶活性分析,一部分做生物碱含量分析,所有数据均为3次平行重复测定的结果,标准偏差不超过10%。
, 百拇医药
1.2 组织生长(率)和酶活性测定:组织生长(率)用鲜重法规定,愈伤组织诱导试验中,直接以收获鲜重表示生长量增长,组织生长率=收获鲜重/接种鲜重×100%。过氧化物酶酶液提取制备和酶活性测定按前文的方法[4],碱性过氧化物酶活性测定在 0.1 mol/L Tris-HCl(pH9.0)缓冲液中进行,酸性过氧化物酶活性测定在 0.05 mol/L 柠檬酸-磷酸氢二钠缓冲液(pH5.5)中进行。
1.3 生物碱的提取和测定:按前文[5]的方法进行,采用外标法测定,以TLC、保留时间、共进样和紫外吸收光谱进行几种生物碱的鉴定。
2 结果与讨论
2.1 长春花叶片愈伤组织诱导发生过程中培养物的生长动态:从生长曲线图1可见,在黑暗和光照条件下,几种诱导培养基均可刺激叶片产生愈伤组织,诱导率均为 100%,在 4 种诱导培养基中,M3 诱导效果最好,愈伤组织浅黄疏松,生长较快,愈伤组织发生较早,生长状态最好。M2 诱导效果其次,愈伤组织发生和生长均很好,但在光下略有褐化现象,使生长受到一些抑制。M1,L1 培养基诱导效果一般,在光照培养中,鲜重增长较慢,褐化严重,长久光照下组织变绿,黑暗中生长良好。光照在早期促进愈伤组织发生和生长,但在后期则对生长有抑制作用。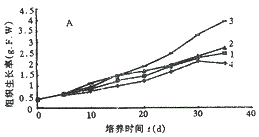

, 百拇医药
A-光照 B-黑暗 1-M1 2-M2 3-M3 4-L1 M1~M3、L1-4 种诱导培养基的培养物(下同)
图1 长春花叶片愈伤组织诱导发生过程中培养物的生长动态

图2 长春花叶片愈伤组织诱导发生过程中培养物的酸性过氧化物酶活性的变化生长动态
图3 长春花叶片愈伤组织诱导发生过程中培养物的碱性过氧化物酶活性的变化动态
2.2 长春花叶片愈伤组织诱导发生过程中酸性和碱性过氧化物酶活性的动态变化:由图 2,图 3 可见,在愈伤组织诱导发生过程中,酸性与碱性过氧化物酶活性基本呈上升趋势。但在 10~20 d 之间不同培养基的培养物的 POD 都有一个峰值,观察结果表明,此峰值时间与愈伤组织发生的时间同步,例如 M3 诱导的愈伤组织发生较早,M3 中培养物便是该峰值提前者,光下比在黑暗中更有利于愈伤组织的发生,体现在峰值较早地出现。2,4-D 明显地抑制 POD 的活性,且在黑暗中的抑制效应更强。无论是酸性还是碱性过氧化物酶活性在光照下都明显高于黑暗中。由图可见,各组织的碱性 POD 在均呈曲折上升。
, http://www.100md.com
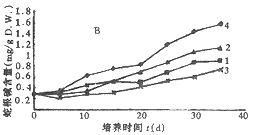
2.3 长春花叶片愈伤组织诱导发生过程中阿玛碱和蛇根碱的动态变化:由图4可见,在植物叶片中含量较低的阿玛碱当叶片愈伤化后开始上升,在L1和M2培养基中上升最快,而在M3中最小,这可能是2,4-D的抑制作用。光照明显降低阿玛碱的积累,在光照条件下阿玛碱只达到0.347 mg/g干重,在黑暗中阿玛碱在L1中可积累至 1.383 mg/g 干重。由图5可见,蛇根碱则同样也受 2,4-D 的抑制,培养物在M3培养基中蛇根碱的含量远比L1中的低,而在光下的L1培养基中蛇根碱则可积累至1.754 mg/g干重。在光照下培养物的蛇根碱总是比黑暗中的含量高,可能是由于光诱导过氧化物酶活性提高的缘故。

图4 长春花叶片愈伤组织诱导发生过程中培养物的阿玛碱含量的变化动态
, 百拇医药
图5 长春花叶片愈伤组织诱导发生过程中培养物的蛇根碱含量的变化动态
2.4 长春花叶片愈伤组织诱导发生过程中文多灵和长春质碱的动态变化:文多灵和长春质碱只在不加2,4-D 的培养基中存在,而且在诱导愈伤组织发生的过程中,其含量发生剧烈变化,离体培养开始文多灵迅速下降,而且降幅最大,在10d内便由1.14mg/g干重降至0.03~0.08 mg/g干重。而长春质碱先上升然后又迅速下降,后期仍保持一定水平(图6,7)。光照有利于缓解文多灵和长春质碱的降解。这两种不同类型生物碱的合成和降解的消长,反映了叶片愈伤组织诱导初期外植体发生的生理生化和次生代谢途径的变化。

图 6 长春花叶片愈伤组织诱导发生过程中培养物的长春质碱含量的变化动态

, 百拇医药
图 7 长春花叶片愈伤组织诱导发生过程中培养物的文多灵含量的变化动态
2.5 生长素和光暗条件对长春花愈伤组织的生物量和生物碱的调节:在不同种类的生长素与 KT 配比的培养基中,愈伤组织随2,4-D 浓度提高,其生物碱含量降低,POD 活性也降低。光照可使酸性和碱性 POD 活性增加,但持续光照使生长受到抑制,同时也使蛇根碱含量增加,而阿玛碱含量下降,这可能是碱性 POD 活性增加的结果。在2,4-D 培养基中加入 IAA 可增加生物碱的含量,将 2,4-D 用 NAA 或 IAA 代替,则生物碱的合成显著增加,过氧化物酶活性增加,而生长率有一定程度的下降(表1)。
表1 不同的生长素和光暗条件对长春花愈伤组织的生物量、生物碱含量和过氧化物酶活性(POD)的影响 培养基
代号
组织生长率
, http://www.100md.com
阿玛碱 蛇根碱 长春质碱
酸性POD 碱性POD
(%)
(mg/g Dry Weight) (ΔOD470/mg protein.min)
A光培养
345.5
0.207
0.525
0.0013
93.75
20.08
, 百拇医药
暗培养
386.1
0.272
0.374
0.0012
52.43
8.25
B光培养
346.23
0.109
0.442
0.0009
68.42
, http://www.100md.com
11.62
暗培养
497.46
0.203
0.219
ND
28.54
10.25
C光培养
400.21
0.057
0.273
ND
, 百拇医药
34.81
17.45
暗培养
528.9
0.125
0.103
ND
14.25
9.0
D光培养
335.26
0.217
0.596
, http://www.100md.com
0.0018
52.10
21.30
暗培养
417.6
0.397
0.399
0.0006
26.38
10.61
E光培养
325.16
0.353
, 百拇医药
0.657
0.0084
125.36
26.53
暗培养
387.36
0.416
0.547
0.0049
72.54
19.26
F光培养
320.64
, http://www.100md.com
0.308
0.623
0.0079
120.63
33.09
暗培养
370.35
0.372
0.564
0.0047
61.75
18.64
注:培养基的激素配比分别是 A: 2,4-D 1 mg/L+KT 0.1 mg/L;B:2,4-D 2 mg/L+KT 0.1 mg/L;C:2,4-D 3 mg/L+KT 0.1 mg/L;D:2,4-D 1 mg/L+IAA 1 mg/L+KT 0.1 mg/L;E:NAA 1 mg/L+IAA 1 mg/L+KT 0.1 mg/L;F:NAA 1 mg/L+KT 0.1 mg/L。ND表示含量甚微,未能检出。
, http://www.100md.com
3 讨论
许多研究结果表明,在由植物体的某一器官诱导形成愈伤组织后,其合成原器官的次生代谢物的能力极大地降低甚至丧失,而与此同时另一些在原外植体中含量低的化合物却在愈伤组织中大量合成,这可能是因为愈伤组织在不同条件下的次生代谢途径改变。本试验证实长春花植物的叶片诱导愈伤组织过程中生物碱的变化便体现了这点,这与其他试验结果相似[6]。本来在长春花叶片中阿玛碱和蛇根碱的含量较低,而文多灵和长春质碱的含量相对较高,但在叶片脱分化形成愈伤组织的过程中,合成文多灵的能力却逐渐丧失,却获得了高水平合成阿玛碱和蛇根碱的能力,这个过程自然涉及多种与发育和分化调节密切相关的基因的表达及酶的调节的事件。因为正是外源激素使叶片组织发生根本变化,而且过氧化物酶被证明参与阿玛碱向蛇根碱的转化和文多灵与长春质碱偶合为长春碱①,故我们在研究外源激素对叶片变为愈伤组织的过程中生物碱合成影响的同时,检测外源激素和光对过氧化物酶的活性的调节,探索此过程中生物碱合成类型的变化与过氧化物酶之间的关系。这对阐明长春花叶片愈伤组织的诱导过程中发生的生物碱代谢类型的变化机制有重要价值,因为这一变化最终导致组织培养的方法不能生产文多灵,长春碱。本文仅是一部分初步的工作,仍需进一步努力探索。
, http://www.100md.com
参考文献
1 Moreno PRH, et al. Plant Cell Tissue and Organ Culture, 1995,42:1
2 Schlatmann JE, et al. Biochnology and Bioengineering, 1993,41:253
3 Shanks JV, et al. Biotechnology and Bioengineering, 1998,58:333
4 赵 剑,等.生物物理学报,1996,12(3):517
5 赵 剑,等.药学学报,1999,34(7):
6 Morris P. Planta Medica, 1986,:127
收稿日期:1998-11-23, 百拇医药(赵 剑* 朱蔚华 胡 秋 王文科)
单位:赵 剑* 朱蔚华 胡 秋 中国协和医科大学药物研究所 北京 100050中国医学科学院;王文科 山西师范大学生物系
关键词:长春花愈伤组织;吲哚生物碱;过氧化物酶
中草药990729 摘 要 在 4 种不同培养基上诱导长春花叶片愈伤组织发生的过程中,叶片中含量较高的文多灵(vindoline)和长春质碱(catharanthine)迅速降解至极低水平,而在叶片中含量较低的阿玛碱(ajmalicine)和蛇根碱(serpentine)的合成却逐渐增加,同时酸性和碱性过氧化物酶(peroxidase)活性呈上升趋势,尤其碱性过氧化物酶与蛇根碱和阿玛碱的相关性较密切。光照和黑暗中的变化趋势基本一致,但光照可提高过氧化物酶的活性,对培养物的长春质碱、文多灵和蛇根碱的合成有利。不同的培养基对生物碱的合成影响较大,2,4-D 显著抑制生物碱的合成,持续光照使愈伤组织的生长受到抑制,使阿玛碱向其蛇根碱转化。
, 百拇医药
Regulation of Indole Alkaloids Synthesis and Related Enzymes During Callus
Initiation and Culture of Madagascar Periwinkle Leaves (Catharanthus roseus)
Zhao Jian, Wang Wenke, Zhu Weihua, et al
(Institute of Materia Medica of PUMC & CAMC, Beijing 100050)
Abstract Leaf calluses of Catharanthus roseus (L.) G.Don were initiated in four different media under illumination or in dark. Visible callus began to form within 10 to 15 days. The ordinarily high contents of vindoline and catharanthine in the leaf declined rapidly, while the lower ajmalicine and serpentine contents increased markedly during callus initiation. At the same time, acidic and alkaline peroxidase showed a tendency to increase their activities, especially the activity of alkaline peroxidase was more closely related with the synthesis of serpentine and ajmalicine. Results also showed that illumination improved occurrence of leaf callus in advance and stimulated peroxidase activity, beneficial for the synthesis of vindoline, catharanthine and serpentine. 2.4-D represses biosynthesis of indole alkaloids and peroxidase activity in all cultures.
, 百拇医药
Key words Catharanthus roseus (L.) G.Don leaf callus initiation indole alkaloid peroxidase activity
长春花组织和细胞培养已经研究了很多年,但至今仍仅限于中等规模的生物反应器生产,且长春花生物碱的生物合成途径和调节有许多不清楚之处,所以国内外的很多实验室仍在进行研究[1~3]。尤其近几年在长春碱(vinblastine)的基础上得到新的半合成抗癌药异长春碱(vinorelbine)具有很大的应用潜力,市场价格高于紫杉醇。而其半合成原料仍需长春质碱和文多灵,使组织培养这一可能的原料来源途径再次受到人们关注。从长春花愈伤组织的诱导、继代培养到细胞悬浮培养这个过程中培养物的生物碱合成代谢类型和能力有了很大变化,这一切是如何发生的还很不清楚,本文试图从愈伤组织的诱导和继代培养过程中的生物碱合成类型的变化和调节及此过程中的相关酶活性变化来探索其原因。
, http://www.100md.com
1 实验材料和方法
1.1 愈伤组织的诱导:取实验室内盆栽的长春花顶芽下第三片叶,用肥皂水和自来水清洗干净,吸干水分,先用70 %酒精灭菌2 min 后,转入0.1 %的升汞灭菌10 min,无菌蒸馏水冲洗5次,剪成小块,接种于以下4种诱导培养基中:M1:MS+NAA 1 mg/L+IAA 1 mg/L+KT 0.1 mg/L+5%蔗糖+0.7%琼脂;M2:MS+NAA 1 mg/L+KT 0.1 mg/L+5%蔗糖+0.7%琼脂;M3:MS+2,4-D 1 mg/L+KT 0.1 mg/L+3%蔗糖+0.7%琼脂;L1:LS+IAA 1 mg/L+6-BA 0.1 mg/L+5%蔗糖+0.7%琼脂,培养基在高温灭菌前均调至pH8.5。每瓶接种物约为0.4 g。分别置于光下和黑暗中培养,培养温度均为 23℃±2℃,光照周期12 h/d,光强约2 000 lx。激素和光的调节实验用来自同一植株的组织系,取继代3次的该愈伤组织均匀地接种于含有不同种类和浓度的生长素和激动素组合的MS培养基含3 %蔗糖和0.7 %琼脂(激素配比编号为 A,B,C,D,E,F),分别置于光下和黑暗中培养,培养基制备和培养条件与以上相同。愈伤组织诱导实验每隔5 d取一次材料,光和激素调节实验则在培养45 d后取材,称鲜重后,一部分做酶活性分析,一部分做生物碱含量分析,所有数据均为3次平行重复测定的结果,标准偏差不超过10%。
, 百拇医药
1.2 组织生长(率)和酶活性测定:组织生长(率)用鲜重法规定,愈伤组织诱导试验中,直接以收获鲜重表示生长量增长,组织生长率=收获鲜重/接种鲜重×100%。过氧化物酶酶液提取制备和酶活性测定按前文的方法[4],碱性过氧化物酶活性测定在 0.1 mol/L Tris-HCl(pH9.0)缓冲液中进行,酸性过氧化物酶活性测定在 0.05 mol/L 柠檬酸-磷酸氢二钠缓冲液(pH5.5)中进行。
1.3 生物碱的提取和测定:按前文[5]的方法进行,采用外标法测定,以TLC、保留时间、共进样和紫外吸收光谱进行几种生物碱的鉴定。
2 结果与讨论
2.1 长春花叶片愈伤组织诱导发生过程中培养物的生长动态:从生长曲线图1可见,在黑暗和光照条件下,几种诱导培养基均可刺激叶片产生愈伤组织,诱导率均为 100%,在 4 种诱导培养基中,M3 诱导效果最好,愈伤组织浅黄疏松,生长较快,愈伤组织发生较早,生长状态最好。M2 诱导效果其次,愈伤组织发生和生长均很好,但在光下略有褐化现象,使生长受到一些抑制。M1,L1 培养基诱导效果一般,在光照培养中,鲜重增长较慢,褐化严重,长久光照下组织变绿,黑暗中生长良好。光照在早期促进愈伤组织发生和生长,但在后期则对生长有抑制作用。
, 百拇医药
A-光照 B-黑暗 1-M1 2-M2 3-M3 4-L1 M1~M3、L1-4 种诱导培养基的培养物(下同)
图1 长春花叶片愈伤组织诱导发生过程中培养物的生长动态
图2 长春花叶片愈伤组织诱导发生过程中培养物的酸性过氧化物酶活性的变化生长动态
图3 长春花叶片愈伤组织诱导发生过程中培养物的碱性过氧化物酶活性的变化动态
2.2 长春花叶片愈伤组织诱导发生过程中酸性和碱性过氧化物酶活性的动态变化:由图 2,图 3 可见,在愈伤组织诱导发生过程中,酸性与碱性过氧化物酶活性基本呈上升趋势。但在 10~20 d 之间不同培养基的培养物的 POD 都有一个峰值,观察结果表明,此峰值时间与愈伤组织发生的时间同步,例如 M3 诱导的愈伤组织发生较早,M3 中培养物便是该峰值提前者,光下比在黑暗中更有利于愈伤组织的发生,体现在峰值较早地出现。2,4-D 明显地抑制 POD 的活性,且在黑暗中的抑制效应更强。无论是酸性还是碱性过氧化物酶活性在光照下都明显高于黑暗中。由图可见,各组织的碱性 POD 在均呈曲折上升。
, http://www.100md.com
2.3 长春花叶片愈伤组织诱导发生过程中阿玛碱和蛇根碱的动态变化:由图4可见,在植物叶片中含量较低的阿玛碱当叶片愈伤化后开始上升,在L1和M2培养基中上升最快,而在M3中最小,这可能是2,4-D的抑制作用。光照明显降低阿玛碱的积累,在光照条件下阿玛碱只达到0.347 mg/g干重,在黑暗中阿玛碱在L1中可积累至 1.383 mg/g 干重。由图5可见,蛇根碱则同样也受 2,4-D 的抑制,培养物在M3培养基中蛇根碱的含量远比L1中的低,而在光下的L1培养基中蛇根碱则可积累至1.754 mg/g干重。在光照下培养物的蛇根碱总是比黑暗中的含量高,可能是由于光诱导过氧化物酶活性提高的缘故。
图4 长春花叶片愈伤组织诱导发生过程中培养物的阿玛碱含量的变化动态
, 百拇医药
图5 长春花叶片愈伤组织诱导发生过程中培养物的蛇根碱含量的变化动态
2.4 长春花叶片愈伤组织诱导发生过程中文多灵和长春质碱的动态变化:文多灵和长春质碱只在不加2,4-D 的培养基中存在,而且在诱导愈伤组织发生的过程中,其含量发生剧烈变化,离体培养开始文多灵迅速下降,而且降幅最大,在10d内便由1.14mg/g干重降至0.03~0.08 mg/g干重。而长春质碱先上升然后又迅速下降,后期仍保持一定水平(图6,7)。光照有利于缓解文多灵和长春质碱的降解。这两种不同类型生物碱的合成和降解的消长,反映了叶片愈伤组织诱导初期外植体发生的生理生化和次生代谢途径的变化。
图 6 长春花叶片愈伤组织诱导发生过程中培养物的长春质碱含量的变化动态
, 百拇医药
图 7 长春花叶片愈伤组织诱导发生过程中培养物的文多灵含量的变化动态
2.5 生长素和光暗条件对长春花愈伤组织的生物量和生物碱的调节:在不同种类的生长素与 KT 配比的培养基中,愈伤组织随2,4-D 浓度提高,其生物碱含量降低,POD 活性也降低。光照可使酸性和碱性 POD 活性增加,但持续光照使生长受到抑制,同时也使蛇根碱含量增加,而阿玛碱含量下降,这可能是碱性 POD 活性增加的结果。在2,4-D 培养基中加入 IAA 可增加生物碱的含量,将 2,4-D 用 NAA 或 IAA 代替,则生物碱的合成显著增加,过氧化物酶活性增加,而生长率有一定程度的下降(表1)。
表1 不同的生长素和光暗条件对长春花愈伤组织的生物量、生物碱含量和过氧化物酶活性(POD)的影响 培养基
代号
组织生长率
, http://www.100md.com
阿玛碱 蛇根碱 长春质碱
酸性POD 碱性POD
(%)
(mg/g Dry Weight) (ΔOD470/mg protein.min)
A光培养
345.5
0.207
0.525
0.0013
93.75
20.08
, 百拇医药
暗培养
386.1
0.272
0.374
0.0012
52.43
8.25
B光培养
346.23
0.109
0.442
0.0009
68.42
, http://www.100md.com
11.62
暗培养
497.46
0.203
0.219
ND
28.54
10.25
C光培养
400.21
0.057
0.273
ND
, 百拇医药
34.81
17.45
暗培养
528.9
0.125
0.103
ND
14.25
9.0
D光培养
335.26
0.217
0.596
, http://www.100md.com
0.0018
52.10
21.30
暗培养
417.6
0.397
0.399
0.0006
26.38
10.61
E光培养
325.16
0.353
, 百拇医药
0.657
0.0084
125.36
26.53
暗培养
387.36
0.416
0.547
0.0049
72.54
19.26
F光培养
320.64
, http://www.100md.com
0.308
0.623
0.0079
120.63
33.09
暗培养
370.35
0.372
0.564
0.0047
61.75
18.64
注:培养基的激素配比分别是 A: 2,4-D 1 mg/L+KT 0.1 mg/L;B:2,4-D 2 mg/L+KT 0.1 mg/L;C:2,4-D 3 mg/L+KT 0.1 mg/L;D:2,4-D 1 mg/L+IAA 1 mg/L+KT 0.1 mg/L;E:NAA 1 mg/L+IAA 1 mg/L+KT 0.1 mg/L;F:NAA 1 mg/L+KT 0.1 mg/L。ND表示含量甚微,未能检出。
, http://www.100md.com
3 讨论
许多研究结果表明,在由植物体的某一器官诱导形成愈伤组织后,其合成原器官的次生代谢物的能力极大地降低甚至丧失,而与此同时另一些在原外植体中含量低的化合物却在愈伤组织中大量合成,这可能是因为愈伤组织在不同条件下的次生代谢途径改变。本试验证实长春花植物的叶片诱导愈伤组织过程中生物碱的变化便体现了这点,这与其他试验结果相似[6]。本来在长春花叶片中阿玛碱和蛇根碱的含量较低,而文多灵和长春质碱的含量相对较高,但在叶片脱分化形成愈伤组织的过程中,合成文多灵的能力却逐渐丧失,却获得了高水平合成阿玛碱和蛇根碱的能力,这个过程自然涉及多种与发育和分化调节密切相关的基因的表达及酶的调节的事件。因为正是外源激素使叶片组织发生根本变化,而且过氧化物酶被证明参与阿玛碱向蛇根碱的转化和文多灵与长春质碱偶合为长春碱①,故我们在研究外源激素对叶片变为愈伤组织的过程中生物碱合成影响的同时,检测外源激素和光对过氧化物酶的活性的调节,探索此过程中生物碱合成类型的变化与过氧化物酶之间的关系。这对阐明长春花叶片愈伤组织的诱导过程中发生的生物碱代谢类型的变化机制有重要价值,因为这一变化最终导致组织培养的方法不能生产文多灵,长春碱。本文仅是一部分初步的工作,仍需进一步努力探索。
, http://www.100md.com
参考文献
1 Moreno PRH, et al. Plant Cell Tissue and Organ Culture, 1995,42:1
2 Schlatmann JE, et al. Biochnology and Bioengineering, 1993,41:253
3 Shanks JV, et al. Biotechnology and Bioengineering, 1998,58:333
4 赵 剑,等.生物物理学报,1996,12(3):517
5 赵 剑,等.药学学报,1999,34(7):
6 Morris P. Planta Medica, 1986,:127
收稿日期:1998-11-23, 百拇医药(赵 剑* 朱蔚华 胡 秋 王文科)