P85宋擻次桓-3爾淡-軌易爾淡C-ζ鹸栽麗斤僮砿諸嫖殆Å爾試峠錆次聾飲p70宰軌易悶S6爾淡議距准恬喘
恬宀⦿僧極啓 Berk BC 購喟坿
汽了⦿僧極啓(財剩匳僥垪伉稽勞尖冩梢片⇧財剩 421001)◉Berk BC(Center for Cardiovascular Research, University of Rochester Medical Center, Rochester, New York 14642, USA)◉購喟坿(嶄表匳親寄僥勞尖僥縮冩片⇧鴻巒 510089)
購囚簡⦿次,峠錆,僮砿◉宋擻次桓-3爾淡◉僮砿諸嫖殆Å◉軌易爾淡C◉p70宰軌易悶S6爾淡
嶄忽強琢啣晒墫崗000302[姜 勣] 恬宀參念議冩梢窟孖軌易爾淡C-ζ初擬僮砿諸嫖殆Å将Ras-MEK余抄爾試僮砿峠錆次聾飲某叢殆試晒議軌易爾淡MAPK賜聾飲翌佚催距准爾淡ERK1/2。云猟冩梢阻P85宋擻次桓-3爾淡-軌易爾淡C-ζ鹸栽麗宰軌易悶爾淡p70S6爾淡試來議距准恬喘式凪佚催勧弓余抄。 Western Blot蛍裂𠮟幣屎械凍劍議僮砿峠錆次聾飲燕器p70S6爾淡、p85宋擻次桓-3爾淡才軌易爾淡C-ζ。100 nmol僮砿諸嫖殆Å才 10 μg/L 僮弌医坿伏海咀徨缶爾旺音唹峒p70S6爾淡、p85宋擻次桓-3爾淡才軌易爾淡C-ζ燕器。 p70S6爾淡宋磨廬卞淡試來蛍裂窟孖僮砿諸嫖殆Å嚥僮砿峠錆次聾飲恬喘5 min 軸蝕兵爾試p70S6爾淡, 20 min器互桁, 旺格質楚卆正來。宋擻次桓-3爾淡雙崙質wortmannin (10 nmol)、軌易爾淡C-ζ 雙崙質Pseudo Z (50 μmol) 才p70S6爾淡雙崙質rapamycin (100 μg/L) 譲辛苧𠮟怦僅僮砿諸嫖殆Å嗾擬議p70S6爾淡爾試。嗤箸議頁⇧厘断鉱賀欺p85宋擻次桓-3爾淡才軌易爾淡C-ζ鞭僮砿諸嫖殆Å才僮弌医坿伏海咀徨缶爾朔譲﨑Ras廬了旺嚥岻潤栽, 森Ras森悶辛繍p85宋擻次桓-3爾淡賜軌易爾淡C-ζ詞栽柿牛, 森p85宋擻次桓-3爾淡森悶呀辛聞軌易爾淡C-ζ柿牛和栖。 戻幣Ras、p85宋擻次桓-3爾淡才軌易爾淡C-ζ眉嶽軌易潤栽壓匯軟侘撹匯倖孔嬬鹸栽悶⇧Wortmannin才Pseudo Z辛怦僅宸倖孔嬬鹸栽悶議侘撹。緩翌⇧僮砿諸嫖殆Å才僮弌医坿伏海咀徨嚥僮砿峠錆次聾飲剣圄24 h 苧𠮟陥序聾飲奐岾⇧覦炎俟寘獨爐用剳宰榠塊秘楕奐紗埃50%, Wortmannin才掲蒙呟來軌易爾淡C雙崙質Chelerythine譲雙崙僮砿諸嫖殆Å才僮弌医坿伏海咀徨斤僮砿峠錆次聾飲議陥奐岾恬喘。忝栽貧峰潤惚, 傍苧僮砿諸嫖殆Å宥狛Ras-宋擻次桓-3爾淡-軌易爾淡C-ζ余抄爾試p70S6爾淡才缶爾僮砿峠錆次聾飲奐岾, p85宋擻次桓-3爾淡-軌易爾淡C-ζ 鹸栽麗壓凪嶄軟嶷勣距准恬喘。
, http://www.100md.com
[嶄夕蛍窃催] Q55 [猟孽炎紛鷹] A
[猟嫗園催] 1007-3949(2000)-03-0193-06
Phosphatidylinositol 3-kinase/Protein Kinase C-ζ Complex Regulates Angiotensin Å Activation of p70 Ribosomal S6 Kinase in Vascular Smooth Muscle Cells
LIAO Duan-Fang
(Division of Cardiopulmonary Pharmacology, Hengyang Medical College, Hengyang 421001, China.)
Bradford C BERK
, 為調匳勞
(Center for Cardiovascular Research, University of Rochester Medical Center, Rochester, New York 14642, USA)
GUAN Yong-Yuan
(Department of Pharmacology, Sun Yat-sen University of Medicine Sciences, Guangzhou 510089)
ABSTRACT Aim To investigate the effects of phosphatidylinositol 3-kinase (PI3K)/protein kinase C-ζ (PKC-ζ)complex on regulating angiotensin Å(AngÅ) activation of p70 ribosomal S6 kinase (p70S6K) in vascular smooth muscle cells(VSMC). Methods VSMC isolated from 200 g to 250 g male Sprague-Dawley rats were cultured to 70% to 80% confluence and were growth-arrested by incubation in 0.1% calf serum/DMEM for 48 h before use. Both 3 H-TdR incorporation and cell counting were used to estimate the proliferation of VSMC. Western blot, immunoprecipitation and immunoblot analyses were performed to valuate the AngÅ-stimulated kinase expression and Ras-PI3K-PKC-ζ association of VSMC. Phosphotransferase assay by using S6 peptide as substrate was employed to measure the p70 S6K activity. Results VSMC expressed p85PI3K, PKC-ζ and p70S6K; AngÅ and PDGF stimulation did not affect the p85PI3K, PKC-ζ and p70S6K expression. 2) 100 nmol AngÅ treatment obviously stimulated p70S6K activity of VSMC at 5 min with peak at 20 min. PI3K inhibitor wortmannin (10 nmol) and pseudosubstrate of PKC-ζ pseudo Z (50 μmol) blocked AngÅ activation of p70S6K. 3) In response to AngÅ, PI3K translocated to the membrane and associated with Ras as shown by PI3K coprecipitation with Ras antibody. Furthermore, both anti-Ras and anti-PI3K antibodies could precipitate PKC-ζ, suggesting that AngÅ treatment resulted in formation of Ras-PI3K-PKC-ζ complex. Wortmannin and pseudo Z significantly blocked the complex formation. 4) In addition, 100 nmol AngÅ and 10 ng/mL PDGF incubation with VSMC for 24 hours stimulated VSMC proliferation, which could be abrogated by wortmannin and nonspecific PKC inhibitor chelerythrine. Conclusion This findings demonstrate that AngÅ stimulation of p70S6K occurs via a pathway of Ras-PI3K-PKC-ζ and p85PI3K-PKC-ζcomplex play an important role in this procedure
, 為調匳勞
MeSH Angiotensin Å◉ Phosphotransferases, Phosphatidylinositol◉ Protein Kinase C◉ Phosphotransferases, RNA Ribosomal S6◉ Muscle, Smooth, Vascular
僮砿諸嫖殆Å(angiotensin Å, AngÅ) 壓僮砿峠錆次聾飲(vascular smooth muscle cells, VSMC)奐岾才景寄狛殻嶄軟嶷勣恬喘[1]。猟孽[2-4]烏祇AngÅ奐紗聾飲坪呼宣徨敵業、爾試軌易爾淡C(protein kinase C, PKC)、試晒匯狼双喇伏海咀徨嗾擬議佚催勧擬宥揃⇧泌爾試軌易籍葦磨爾淡、爾試某叢殆試晒議軌易爾淡(mitogine activated-protein kinase, MAPK)賜聾飲翌佚催距准爾淡(extracelluar signal-regulated kinases 1/2, ERK1/2) 吉吉。P70宰軌易悶爾淡(p70 ribosomal S6 kinase, p70S6爾淡)狼某葦磨/釦葦磨軌易爾淡社怛撹埀。輝聾飲歌嚥斤伏海咀徨、蛍叢吉壼豚郡哘扮⇧p70S6爾淡瓜爾試。猟孽烏祇壓白漬鱒彬議寄報麼強琢才伉縫伉壜⇧p70S6爾淡瓜儻堀爾試[1,5]。P70S6K宥狛距准mRNA議鍬咎狛殻糞孖斤聾飲伏海議距陣[6,7], p70S6爾淡議試晒鞭凪貧嗄宋擻次桓-3爾淡(phosphatidylinositol 3-kinse, PI3K)議距准[8]。猟孽[9,10]烏祇AngÅ 辛缶爾凍劍議VSMC才伉次聾飲 p70S6爾淡試來, 隼遇凪佚催勧擬余抄旺音賠萱⇧容霞嚥Ras-Raf-MEK-ERK1/2嗤購。恬宀參念議垢恬𠮟幣Raf 壓AngÅ 爾試ERK1/2狛殻嶄旺音軟麼勣恬喘⇧遇喇PKC-ζ函遇旗岻[11,12]。Saward 吉[13]烏祇1 μmol AngÅ嚥VSMC恬喘15 min辛爾試PI3K⇧聞PI3K p85冉汽了籍葦磨宋磨晒旺窟伏弔廬了。除定音富猟孽𠮟幣PKC-ζ頁PI3K議和嗄丼哘軌易, 揖扮匆歌嚥p70S6爾淡議距准, 凪試來鞭PI3K旗仍恢麗PtdIns(3-5)P(3)議距陣[14-16]。咀遇⇧厘断容霞PKC-ζ呀辛嬬初擬AngÅ爾試p70S6爾淡, 旺戻竃PI3K-PKC-ζ選栽距陣p70S6爾淡試來議垢恬邪譜。
, 為調匳勞
1 可創嚥圭隈
1.1 可創
森H-Ras森悶才森PKC-ζ森悶蛍艶杭徭Boehringer Mannheim才Santa Cruz巷望⇧森p70S6爾淡森悶才森p85PI3K森悶杭徭Upstate Biotechnology Inc巷望⇧S6訥喇紗鎮寄Kinetek Pharmaceuticals Inc巷望奘僕⇧[r-32P]ATP葎Dupont巷望恢瞳⇧Protein A-agarose 葎Life Technologies Inc巷望恢瞳⇧wortmannin、chelerythrine才rapamycin 葎Sigma巷望恢瞳⇧PKC-ζ邪來久麗杭徭Calbiochem巷望(La Jolla, CA)。
1.2 僮砿峠錆次聾飲凍劍
歌孚猟孽[17]⇧貫200゛250 g俛來Sprague Dawley寄報俟麼強琢蛍宣竃VSMC⇧喘根10%弌釘僮賠議DMEM凍劍児凍劍。函5゛13旗VSMC噐100 cm凍劍駐嶄凍劍崛70%゛80%蛮栽⇧喘根0.1%弌釘僮賠議DMEM凍劍児写偬凍劍48 h聞VSMC侃噐床峭彜蓑⇧隼朔喘AngÅ、 PDGF吉缶爾20 min。PI3K議雙崙質Wortmnnin、掲蒙呟來PKC-雙崙質Chelerythine、PKC-ζ 雙崙質(PKC-ζ邪來久麗)Pseudo Z才p70S6爾淡雙崙質rapamycin噐AngÅ、PDGF岻念10 min紗秘。
, http://www.100md.com
1.3 聾飲軌易戻函嚥敵業霞協
僮砿諸嫖殆Å(AngÅ)、 PDGF恬喘朔議VSMC喘宋磨産喝匣牢眉肝⇧耽駐紗1.0 mL TME産喝匣(10 mmol Tris, 5 mmol MgCl2, 1 mmol EDTA, 25 mmol NaF, pH 7.5◉匝喘念紗100 μmol Na3VO4, 20 mg/L leupeptin, 1 mg/L pepstatin A, 4 mg/L aprotinin, 1 mmol DTT) 叢盾聾飲。堀恭, 甥圈貧産蛸鹸釦⇧肯和聾飲⇧甥圈貧喘階蕗頚哩聾飲⇧15 000g宣伉30 min, 函貧賠匣⇧梓Bradford軌易霞協歳傍苧慕霞協軌易敵業⇧劔瞳廈贋噐-80≧棋喘。
1.4 p70S6爾淡試來霞協
歌孚猟孽[18]⇧函森p70S6爾淡森悶窒吽柿牛麗(50% bead slurry) 5 μL, 紗30 μL郡哘産喝匣(25 mmol b-glycerophosphate, 20 mmol MOPS, pH 7.2, 5 mmol EGTA, 2 mmol EDTA, 20 mmol MgCl2, 1 mmol Na3VO4, 0.25 mmol dithiothreitol), 紗4 μg 久麗S6膏訥才50 μmol [r-32 P]ATP(2〜1015 dpm/mol), 噐30≧郡哘15 min, 函20 μL郡哘詞栽麗泣劔噐2 cm2 p81 phosphocellulose 郡哘崕(Whatman巷望)⇧喘根1% orthophosphoric acid 肇宣徨邦喝牢眉肝⇧喘r-柴方匂芝村慧符試來。
, http://www.100md.com
1.5 Western Blot 糞刮
歌孚猟孽[11]⇧塘崙9%議SDS-PAGE蛍宣什才3%議持蚊什⇧耽迅紗秘20 μg軌易劔瞳⇧崔窮咯産喝匣嶄⇧70 V窮咯埃20 min。棋劔瞳序秘蛍宣什朔⇧150 V窮咯崛侭俶扮寂。 繍軌易廬咫崛嵓磨亘略院弔⇧喘Life Technologies巷望斌喘撃液匣噐片梁侃尖2 h⇧梓1|200紗p70S6爾淡、p85PI3K賜PKC-ζ汽森, 4≧狛匚⇧TBS匣牢弔10 min〜3肝⇧喘星功狛剳晒麗淡編質歳(ECL, Amersham International plc. United Kingdom)𠮟弼。
1.6 Ras-宋擻次桓-3爾淡-軌易爾淡C-ζ詞栽柿牛糞刮
歌孚猟孽[12]⇧函TME産喝匣戻函議軌易⇧紗森Ras汽森賜p85PI3K汽森才protein A-agarose噐4≧詞栽12 h序佩窒吽柿牛(immunoprecipitation, IP)。 Protein A-agarose喘TME産喝匣牢匯肝⇧TTBS産喝匣(20 mmol Tris, pH 7.5, 500 mmol NaCl, 1% Triton X-100, 0.1% beta-mercaptoethanol)牢屈肝⇧ 壅喘TME産喝匣牢匯肝。隼朔⇧喘9%議SDS-PAGE什繍窒吽鹸栽麗蛍宣⇧ 喘森p85PI3K才PKC-ζ汽森序佩窒吽咫治(immunoblot, IB)糞刮。
, 為調匳勞
1.7 聾飲柴方嚥覦炎俟寘獨爐用剳宰榠塊秘糞刮
歌孚猟孽[13]⇧函伏海措挫議VSMC崙撹聾飲傅匣(4〜107倖/L)俊嶽噐24迅凍劍医嶄⇧耽迅0.5 mL(2〜104倖聾飲)⇧剣圄24 h。算涙僮賠凍劍児写偬剣圄24 h朔⇧聞VSMC侃噐床峭豚⇧紗弌釘僮賠崛敵業葎10%⇧揖扮蛍怏紗秘100 nmol AngÅ(嶮敵業,和揖) 賜10 μg/L PDGF(嶮敵業,和揖)。 Wortmannin (嶮敵業10 nmol,和揖)、chelerythrine (嶮敵業10 μmol,和揖)才rapamycin(嶮敵業100 nmol,和揖)噐AngÅ、PDGF念10 min 紗秘⇧斤孚怏葎峪根10%弌釘僮賠議DMEM凍劍児, 写偬凍劍24 h。隼朔序佩和双糞刮: 〙聾飲嶧晒匣蛍宣聾飲⇧參4%岬杜清半弼柴方。〖 写偬凍劍18 h, 紗覦炎俟寘獨爐用剳宰榠(3 H-thymine deoxyribonucleotide, 3 H-TdR)崛嶮敵業葎2 mCi/L, 6 h朔辺劔⇧虹根3 H-TdR凍劍児⇧喘D-hanks匣牢眉肝⇧參聾飲嶧晒匣嶧晒。喘寔腎叡繍聾飲渇簾噐横詮亘略陀崕貧⇧5%眉柁閑磨耕協⇧涙邦厰桓用邦⇧孤夬朔崔描妨匿嶄紗描妨匣(遮噂500 mL, PPO 3, POPOP 0.1 g)噐匣悶描妨蛍裂卩(Packard 巷望 )霞慧符來膿業⇧參耽蛍嶝縫受方(decay per minute, dpm賜d/min)燕幣。
, http://www.100md.com
1.8 由柴侃尖
方象參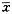 \s燕幣⇧方象将圭餓蛍裂朔序佩q殊刮⇧餓呟議𠮟广來喘P<0.05才P<0.01登僅。
\s燕幣⇧方象将圭餓蛍裂朔序佩q殊刮⇧餓呟議𠮟广來喘P<0.05才P<0.01登僅。
2 潤 惚
2.1 僮砿峠錆次聾飲燕器p70宰軌易悶S6爾淡、p85宋擻次桓-3爾淡才軌易爾淡C-ζ
Western Blot 蛍裂窟孖屎械凍劍議VSMC燕器p70S6爾淡、p85PI3K冉汽了才PKC-ζ, AngÅ才 PDGF缶爾旺音唹峒p70S6爾淡、p85PI3K冉汽了才PKC-ζ議燕器(夕1, Figure 1)。
夕1 僮砿峠錆次聾飲燕器p70宰軌易悶S6爾淡、p85宋擻次桓-3爾淡冉侏才軌易爾淡C-ζ
, http://www.100md.com
Figure 1 VSMC express p70S6K, p85PI3K subtype and PKC-ζ Western blot analysis was carried out with anti- p70S6K antibody (A), anti-p85PI3K antibody (B) and anti- p70S6K, antibody (C), respectively.
2.2 僮砿諸嫖殆Å爾試僮砿峠錆次聾飲p70宰軌易悶S6爾淡
夕2 (Figure 2) 𠮟幣AngÅ嚥VSMC恬喘5 min軸蝕兵爾試p70S6爾淡, 20 min器互桁⇧埃葎侃尖念議5蔚⇧昧朔幟愁週詰(A), 旺格質楚卆正來(B)。p70S6爾淡雙崙質rapamycin (100 nmol)、PI3K雙崙質wortmannin(10 nmol)才PKC-ζ雙崙質Pseudo Z (50 μmol)譲辛嗤丼怦僅AngÅ議恬喘(夕3, Figure 3)。
, 為調匳勞
2.3 僮砿諸嫖殆Å缶爾Ras-宋擻次桓-3爾淡-軌易爾淡C-ζ鹸栽麗侘撹
聾飲鞭AngÅ才 PDGF缶爾朔⇧辛鉱賀欺PI3K﨑Ras廬了旺嚥岻潤栽議孖嵆, 壓森Ras森悶柿牛議軌易嶄嗤PI3K贋壓。夕3A(Figure 3A)嶄辛需森Ras森悶柿牛議軌易窮咯蛍宣廬弔朔壅喘森PI3K森悶墫住辛𠮟幣p85訳揮⇧ wortmannin 10 nmol辛怦僅Ras-PI3K屢札潤栽。夕4B才夕4C(Figure 4B and 4C)嶄辛需音徽森Ras森悶辛繍PKC-ζ詞栽柿牛(B); 遇拝森PI3K森悶匆辛繍PKC-ζ柿牛和栖, PI3K、PKC-ζ岻寂議屢札恬喘呀辛瓜wortmannin才pseudo Z怦僅(C)。喇緩⇧厘断辛參容僅聾飲鞭AngÅ才 PDGF缶爾朔, Ras、PI3K、PKC-ζ眉嶽軌易潤栽壓匯軟侘撹匯倖孔嬬鹸栽悶。Wortmannin才pseudo Z辛怦僅宸倖孔嬬鹸栽悶議侘撹, 傍苧PI3K-PKC-ζ壓宸倖孔嬬鹸栽悶嶄軟嶷勣恬喘。
, 為調匳勞
夕2 僮砿諸嫖殆Å斤僮砿峠錆次聾飲p70宰軌易悶S6爾淡試來議唹峒
Figure 2 The effects of AngÅ on p70S6K activity of VSMC A⦿ Time-course of AngÅ on p70S6K activity. B⦿ Concentration-response of AngÅ on p70S6K activity.
夕3 p70宰軌易悶S6爾淡雙崙質rapamycin、宋擻次桓-3爾淡雙崙質wortmannin才軌易爾淡C-ζ雙崙質pseudo Z斤僮砿諸嫖殆Å試晒僮砿峠錆次聾飲p70宰軌易悶S6爾淡議唹峒
Figure 3 The effects of p70S6K inhibitor rapamycin⇧PI3K inhibitor wortmannin and PKC-ζ inhibitor pseudo Z on AngÅ activation of p70S6K in VSMC a⦿ P<0.01 compared with control grpip, b: P<0.01 compared with AngÅ group, c: P<0.01 compared with PDGF group.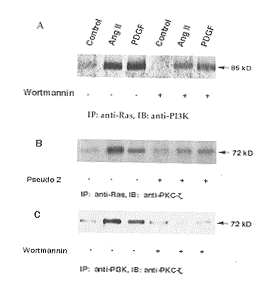
, 為調匳勞
夕4 p85宋擻次桓-3爾淡才軌易爾淡C嚥Ras議選狼
Figure 4 Ras association with p85PI3K and PKC-ζ A and B. p85PI3K and PKC-ζ were immuno-precipitated (IP) with Ras antibody. C. PKC-ζ was immuno-precipitated (IP) by p85PI3K antibody.
2.4 Wortmannin才chelerythrine怦僅僮砿諸嫖殆Å嗾擬僮砿峠錆次聾飲奐岾
岬杜清半弼聾飲柴方才3 H-TdR塊秘隈蛍裂𠮟幣AngÅ(100 nmol)才PDGF (10 μg/L)譲嗤熟膿議缶爾VSMC奐岾議恬喘⇧AngÅ才PDGF侃尖怏聾飲柴方苧𠮟互噐斤孚怏⇧3 H-TdR塊秘楚𠮟广奐紗。Wortmannin 才chelerythrine 譲辛怦僅AngÅ才PDGF議恬喘(燕1, Table 1)。
, http://www.100md.com
燕1 僮砿諸嫖殆Å、僮弌医坿伏海咀徨、Wortmannin才chelerythrine斤僮砿峠錆次聾飲奐岾議唹峒
Table 1 Effects of Ang Å⇧ PDGF⇧ wortmannin and chelerythrine on VSMC proliferation (n=6,\s) Groups
Cell number
(〜104)
3 H-TdR incorporation
(dpm)
Control
, 為調匳勞
2.26\0.14
503\51
AngÅ
3.32\0.29a
740\75a
PDGF
3.51\0.46a
754\95a
Wortmannin
2.25\0.21
512\41
, 為調匳勞
AngÅ+Wortmannin
2.32\0.12b
540\34b
PDGF+Wortmannin
2.31\0.23c
542\60c
Chelerythrine
2.42\0.43
493\75
AngÅ+Chelerythrine
, 為調匳勞
2.47\0.43b
550\84b
PDGF+Chelerythrine
2.42\0.36c
560\71c
a:P<0.01 vs control; b: P<0.01 vs Ang Å◉ c: P<0.01 vs PDGF
3 網 胎
云猟議麼勣窟孖頁PKC-ζ才PI3K選栽距准AngÅ爾試僮砿峠錆次聾飲宰軌易悶爾淡p70S6爾淡。嗤泌和曾泣屶隔宸匯潤胎:〙AngÅ嚥VSMC恬喘5 min 軸蝕兵爾試p70S6爾淡, 20 min器互桁, 旺格質楚卆正來。P70S6K雙崙質rapamycin、 PI3K雙崙質wortmannin才PKC-ζ雙崙質(PKC邪來久麗) Pseudo Z譲辛苧𠮟怦僅AngÅ嗾擬議p70S6爾淡爾試◉ 〖 聾飲鞭AngÅ才 PDGF缶爾朔⇧侘撹Ras-PI3K-PKC-ζ孔嬬鹸栽悶⇧wortmannin才chelerythrine辛雙崙宸倖孔嬬鹸栽悶議侘撹。參念範葎AngÅ麼勣哈軟VSMC景寄, 除定窟孖AngÅ匆醤嗤伏海咀徨議蒙來⇧辛哈軟聾飲奐岾[1,13]。厮岑伏海咀徨宥狛凪聾飲弔貧議軌易籍葦磨鞭悶⇧繍佚催勧公Ras, Ras窟伏弔廬了旺爾試某葦磨/釦葦磨爾淡Raf, 朔宀頁軌易籍葦磨爾淡鞭悶佚催勧擬議嶄伉⇧Raf宥狛MEK爾試ERK1/2序佩佚催選雫慧寄⇧ERK1/2 (嗽各某叢殆爾試議爾淡, mitogen-activated protein kinases, MAPK)繍佚催勧崛聾飲宰⇧尼強DNA栽撹才屢購軌易栽撹, 陥序VSMC奐岾。隼遇⇧AngÅ 旺音恬喘噐軌易籍葦磨鞭悶, 遇頁恬喘噐7-TM鞭悶(seven transmembrane receptors, 7-TM)。7-TM鞭悶頁匯嶽G-軌易鞭悶, 旺音醤嗤籍葦磨爾淡試來。椎担⇧AngÅ頁奕劔陥序聾飲奐岾議椿? 音富猟孽烏祇戻幣軌易籍葦磨鞭悶嚥7-TM鞭悶岻寂贋壓屢札"斤三"(cross talk)[19゛21]。AngÅ辛処祇伏海咀徨佚催勧弓宥揃。猟孽[21⇧22]烏祇AngÅ辛爾試聾飲坪謹嶽軌易爾淡式凪佚催宥祇⇧泌Raf、PKC、PI3K、PLC吉吉。匯違範葎AngÅ辛宥狛曾訳余抄爾試p70S6爾淡: 匯訳辛嬬宥狛Ras-Raf-MEK-ERK1/2余抄。恬宀參念議冩梢窟孖Raf壓AngÅ爾試ERK1/2嶄旺音軟麼勣恬喘⇧遇喇PKC-ζ 函遇旗岻[11,12]。総匯訳辛嬬宥狛Ras-PI3K余抄[8゛10,12]⇧隼遇⇧凪峋聾宥揃旺音賠萱。 猟孽[14゛16]烏祇PKC-ζ歌嚥p70S6爾淡議距准旺初擬凪貧嗄PI3K議丼哘⇧PKC-ζ議試來卆正噐PI3K議恢麗PIP3。厘断容霞PKC-ζ才PI3K選栽歌嚥AngÅ爾試p70S6爾淡議距准。云糞刮鉱賀欺VSMC單米燕器p85PI3K才PKC-ζ, AngÅ質楚卆正來仇爾試p70S6爾淡。 Rapamycin、wortmannin才Pseudo Z譲辛怦僅AngÅ議恬喘。嗤箸議頁⇧厘断窟孖聾飲鞭AngÅ才 PDGF缶爾朔⇧侘撹Ras-PI3K-PKC-ζ孔嬬鹸栽悶⇧森Ras森悶才森PI3K森悶辛詞栽柿牛PKC-ζ, wortmannin才pseudo Z譲辛怦僅緩鹸栽悶議侘撹。緩翌⇧厘断喘VSMC奐岾恬葎嶮泣峺炎蛍裂阻PI3K才PKC-ζ議恬喘⇧ 鉱賀欺PI3K雙崙質wortmannin才掲蒙呟來PKC雙崙質chelerythrine譲辛苧𠮟雙崙AngÅ嗾擬議VSMC奐岾。潤栽參念議垢恬⇧厘断戻竃AngÅ爾試僮砿峠錆次聾飲p70S6爾淡辛嬬淫根泌和佚催勧擬余抄⦿AngÅ 恬喘噐VSMC AT1鞭悶⇧爾試PI3K, 試晒議PI3K嗾擬PKC-ζ匯揖嚥Ras潤栽侘撹孔嬬鹸栽悶, 旺爾試PKC-ζ, 恷嶮爾試p70S6爾淡, 擬崑VSMC奐岾(夕5, Figure 5)。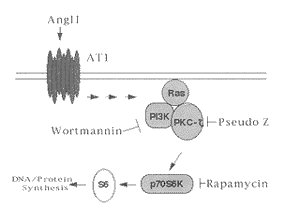
, 為調匳勞
夕5 僮砿諸嫖殆Å将喇Ras-宋淡次桓-3爾淡-軌易爾淡C-ζ爾試p70宰軌易悶S6爾淡議余抄
Figure 5 AngÅ activates p70S6K via the Ras-PI3K-PKC-ζ pathway.
[崑仍] 云猟何蛍垢恬壓胆忽鯖腹禽寄僥Dr. Berk糞刮片才紗鎮寄悟戴曳冉寄僥Dr. Pelech糞刮片頼撹。
[児署𡸴朕] 忽社徭隼親僥児署彿廁仁籾(39970847)
[恬宀酒初] 僧極啓⇧槻⇧刷掴福邃臭偏繁⇧1959定竃伏⇧鴬平冩梢伏。購喟坿⇧槻⇧鴻叫福繁⇧嶄表匳親寄僥勞尖僥縮娩⇧鴬平冩梢伏擬弗⇧忽暦垪僥了溜埀氏勞僥親得咏怏撹埀⇧ゞ嶄忽勞尖僥宥烏〃械暦園溜⇧嶄忽勞尖僥氏伉僮砿廨匍溜埀氏械暦溜埀。
歌深猟孽
, http://www.100md.com
[1] Berk BC. Angiotensin Å signal transduction in vascular smooth muscle: pathways activated by specific tyrosine kinases [J]. J Am Soc Nephrol⇧ 1999⇧ 10: S62-S68
[2] Taubman MB, Berk BC, Izumo S, et al. Angiotensin Å induces c-fos mRNA in aortic smooth muscle cells: role of Ca2+ mobilization and protein kinase C activation [J]. J Biol Chem, 1989, 264: 526-530
[3] Molloy CJ, Taylor DS, Weber H. Angiotensin Å stimulation of rapid protein tyrosine phosphorylation and protein kinase activation in rat aortic smooth muscle cells [J]. J Biol Chem, 1993, 268 (10): 7 338-345
, 為調匳勞
[4] Duff JL, Berk BC, Corson MA. Angiotensin Å stimulates the pp44 and pp42 mitogen-activated protein kinases in cultured rat aortic smooth muscle cells [J]. Biochem Biophys Res Commun, 1992, 188: 257-264
[5] Griendling KK, Ushio-Fukai M, Lassegue B, et al. Angiotensin Å signaling in vascular smooth muscle. New concepts [J]. Hypertension, 1997, 29 (1 Pt 2): 366-373
[6] Pullen N, Thomas G. The modular phosphorylation and activation of p70S6k [J]. FEBS Lett, 1997, 410 (1): 78-82
, 為調匳勞
[7] Avruch J. Insulin signal transduction through protein kinase cascades [J]. Mol Cell Biochem, 1998, 182 (1-2): 31-48
[8] Chung J, Grammer TC, Lemon KP, et al. PDGF- and insulin-dependent pp70S6k activation mediated by phosphatidylinositol-3-OH kinase [J]. Nature, 1994, 370 (6484): 71-75
[9] Giasson E, Meloche S. Role of p70S6 protein kinase in angiotensin Å-induced protein synthesis in vascular smooth muscle cells [J]. J Biol Chem, 1995, 270 (10): 5 225-231
, 為調匳勞
[10] Sadoshima J, Izumo S. Rapamycin selectively inhibits angiotensin Å-induced increase in protein synthesis in cardiac myocytes in vitro. Potential role of 70-kD S6 kinase in angiotensin Å-induced cardiac hypertrophy [J]. Circ Res, 1995, 77 (6): 1 040-052
[11] Liao DF, Duff JL, Daum G, et al. Angiotension Å stimulates MAP kinase kinase kinase activity in vascular smooth muscle cell. Role of Raf [J]. Circ Res, 1996, 79 (5): 1 007-014
, 為調匳勞
[12] Liao DF, Monia B, Dean N, et al. Protein kinase C-ζ mediates angiotensin Å activation of ERK1/2 in vascular smooth muscle cells [J]. J Biol Chem, 1997, 272: 6 146-150
[13] Saward L, Zahradka P. Angiotensin Å activates phosphatidylinositol 3-kinase in vascular smooth muscle cells [J]. Circ Res, 1997, 81 (2): 249-257
[14] Chou MM, Hou W, Johnson J, et al. Regulation of protein kinase C zeta by PI3-kinase and PDK-1 [J]. Curr Biol, 1998, 8 (19): 1 069-077
, 為調匳勞
[15] Le Good JA, Ziegler WH, Parekh DB, et al. Protein kinase C isotypes controlled by phosphoinositide 3-kinase through the protein kinase PDK1 [J]. Science, 1998, 281 (5385): 2 042-045
[16] Romanelli A, Martin KA, Toker A, et al. p70S6 kinase is regulated by protein kinase C zeta and participates in a phosphoinositide 3-kinase-regulated signalling complex [J]. Mol Cell Biol, 1999, 19 (4): 2 921-928
[17] Liao DF, Jin ZG, Baas AS, et al. Purification and identification of secreted oxidative stress-induced factors from vascular smooth muscle cells [J]. J Biol Chem, 2000, 275 (1): 189-196
, 為調匳勞
[18] Xu YJ, Ouk Kim S, Liao DF, et al. Stimulation of 90- and 70-kDa ribosomal protein S6 kinases by arginine vasopressin and lysophosphatidic acid in rat cardiomyocytes [J]. Biochem Pharmacol, 2000, 59 (9): 1 163-171
[19] Velloso LA, Folli F, Sun XJ, et al. Cross-talk between the insulin and angiotensin signaling systems [J]. Proc Natl Acad Sci USA, 1996, 93 (22): 12 490-495
[20] Eguchi S, Numaguchi K, Iwasaki H, et al. Calcium-dependent epidermal growth factor receptor transactivation mediates the angiotensin Å-induced mitogen-activated protein kinase activation in vascular smooth muscle cells [J]. J Biol Chem, 1998, 273 (15): 8 890-896
, 為調匳勞
[21] Berk BC, Corson MA. Angiotensin Å signal transduction in vascular smooth muscle: role of tyrosine kinases [J]. Circ Res, 1997, 80 (5): 607-616
[22] Eguchi S, Iwasaki H, Ueno H, et al. Intracellular signaling of angiotensin Å-induced p70S6 kinase phosphorylation at Ser(411) in vascular smooth muscle cells. Possible requirement of epidermal growth factor receptor, Ras, extracellular signal-regulated kinase, and Akt [J]. J Biol Chem, 1999, 274 (52): 36 843-851
(緩猟2000-07-11辺欺, 2000-09-05俐指), http://www.100md.com
汽了⦿僧極啓(財剩匳僥垪伉稽勞尖冩梢片⇧財剩 421001)◉Berk BC(Center for Cardiovascular Research, University of Rochester Medical Center, Rochester, New York 14642, USA)◉購喟坿(嶄表匳親寄僥勞尖僥縮冩片⇧鴻巒 510089)
購囚簡⦿次,峠錆,僮砿◉宋擻次桓-3爾淡◉僮砿諸嫖殆Å◉軌易爾淡C◉p70宰軌易悶S6爾淡
嶄忽強琢啣晒墫崗000302[姜 勣] 恬宀參念議冩梢窟孖軌易爾淡C-ζ初擬僮砿諸嫖殆Å将Ras-MEK余抄爾試僮砿峠錆次聾飲某叢殆試晒議軌易爾淡MAPK賜聾飲翌佚催距准爾淡ERK1/2。云猟冩梢阻P85宋擻次桓-3爾淡-軌易爾淡C-ζ鹸栽麗宰軌易悶爾淡p70S6爾淡試來議距准恬喘式凪佚催勧弓余抄。 Western Blot蛍裂𠮟幣屎械凍劍議僮砿峠錆次聾飲燕器p70S6爾淡、p85宋擻次桓-3爾淡才軌易爾淡C-ζ。100 nmol僮砿諸嫖殆Å才 10 μg/L 僮弌医坿伏海咀徨缶爾旺音唹峒p70S6爾淡、p85宋擻次桓-3爾淡才軌易爾淡C-ζ燕器。 p70S6爾淡宋磨廬卞淡試來蛍裂窟孖僮砿諸嫖殆Å嚥僮砿峠錆次聾飲恬喘5 min 軸蝕兵爾試p70S6爾淡, 20 min器互桁, 旺格質楚卆正來。宋擻次桓-3爾淡雙崙質wortmannin (10 nmol)、軌易爾淡C-ζ 雙崙質Pseudo Z (50 μmol) 才p70S6爾淡雙崙質rapamycin (100 μg/L) 譲辛苧𠮟怦僅僮砿諸嫖殆Å嗾擬議p70S6爾淡爾試。嗤箸議頁⇧厘断鉱賀欺p85宋擻次桓-3爾淡才軌易爾淡C-ζ鞭僮砿諸嫖殆Å才僮弌医坿伏海咀徨缶爾朔譲﨑Ras廬了旺嚥岻潤栽, 森Ras森悶辛繍p85宋擻次桓-3爾淡賜軌易爾淡C-ζ詞栽柿牛, 森p85宋擻次桓-3爾淡森悶呀辛聞軌易爾淡C-ζ柿牛和栖。 戻幣Ras、p85宋擻次桓-3爾淡才軌易爾淡C-ζ眉嶽軌易潤栽壓匯軟侘撹匯倖孔嬬鹸栽悶⇧Wortmannin才Pseudo Z辛怦僅宸倖孔嬬鹸栽悶議侘撹。緩翌⇧僮砿諸嫖殆Å才僮弌医坿伏海咀徨嚥僮砿峠錆次聾飲剣圄24 h 苧𠮟陥序聾飲奐岾⇧覦炎俟寘獨爐用剳宰榠塊秘楕奐紗埃50%, Wortmannin才掲蒙呟來軌易爾淡C雙崙質Chelerythine譲雙崙僮砿諸嫖殆Å才僮弌医坿伏海咀徨斤僮砿峠錆次聾飲議陥奐岾恬喘。忝栽貧峰潤惚, 傍苧僮砿諸嫖殆Å宥狛Ras-宋擻次桓-3爾淡-軌易爾淡C-ζ余抄爾試p70S6爾淡才缶爾僮砿峠錆次聾飲奐岾, p85宋擻次桓-3爾淡-軌易爾淡C-ζ 鹸栽麗壓凪嶄軟嶷勣距准恬喘。
, http://www.100md.com
[嶄夕蛍窃催] Q55 [猟孽炎紛鷹] A
[猟嫗園催] 1007-3949(2000)-03-0193-06
Phosphatidylinositol 3-kinase/Protein Kinase C-ζ Complex Regulates Angiotensin Å Activation of p70 Ribosomal S6 Kinase in Vascular Smooth Muscle Cells
LIAO Duan-Fang
(Division of Cardiopulmonary Pharmacology, Hengyang Medical College, Hengyang 421001, China.)
Bradford C BERK
, 為調匳勞
(Center for Cardiovascular Research, University of Rochester Medical Center, Rochester, New York 14642, USA)
GUAN Yong-Yuan
(Department of Pharmacology, Sun Yat-sen University of Medicine Sciences, Guangzhou 510089)
ABSTRACT Aim To investigate the effects of phosphatidylinositol 3-kinase (PI3K)/protein kinase C-ζ (PKC-ζ)complex on regulating angiotensin Å(AngÅ) activation of p70 ribosomal S6 kinase (p70S6K) in vascular smooth muscle cells(VSMC). Methods VSMC isolated from 200 g to 250 g male Sprague-Dawley rats were cultured to 70% to 80% confluence and were growth-arrested by incubation in 0.1% calf serum/DMEM for 48 h before use. Both 3 H-TdR incorporation and cell counting were used to estimate the proliferation of VSMC. Western blot, immunoprecipitation and immunoblot analyses were performed to valuate the AngÅ-stimulated kinase expression and Ras-PI3K-PKC-ζ association of VSMC. Phosphotransferase assay by using S6 peptide as substrate was employed to measure the p70 S6K activity. Results VSMC expressed p85PI3K, PKC-ζ and p70S6K; AngÅ and PDGF stimulation did not affect the p85PI3K, PKC-ζ and p70S6K expression. 2) 100 nmol AngÅ treatment obviously stimulated p70S6K activity of VSMC at 5 min with peak at 20 min. PI3K inhibitor wortmannin (10 nmol) and pseudosubstrate of PKC-ζ pseudo Z (50 μmol) blocked AngÅ activation of p70S6K. 3) In response to AngÅ, PI3K translocated to the membrane and associated with Ras as shown by PI3K coprecipitation with Ras antibody. Furthermore, both anti-Ras and anti-PI3K antibodies could precipitate PKC-ζ, suggesting that AngÅ treatment resulted in formation of Ras-PI3K-PKC-ζ complex. Wortmannin and pseudo Z significantly blocked the complex formation. 4) In addition, 100 nmol AngÅ and 10 ng/mL PDGF incubation with VSMC for 24 hours stimulated VSMC proliferation, which could be abrogated by wortmannin and nonspecific PKC inhibitor chelerythrine. Conclusion This findings demonstrate that AngÅ stimulation of p70S6K occurs via a pathway of Ras-PI3K-PKC-ζ and p85PI3K-PKC-ζcomplex play an important role in this procedure
, 為調匳勞
MeSH Angiotensin Å◉ Phosphotransferases, Phosphatidylinositol◉ Protein Kinase C◉ Phosphotransferases, RNA Ribosomal S6◉ Muscle, Smooth, Vascular
僮砿諸嫖殆Å(angiotensin Å, AngÅ) 壓僮砿峠錆次聾飲(vascular smooth muscle cells, VSMC)奐岾才景寄狛殻嶄軟嶷勣恬喘[1]。猟孽[2-4]烏祇AngÅ奐紗聾飲坪呼宣徨敵業、爾試軌易爾淡C(protein kinase C, PKC)、試晒匯狼双喇伏海咀徨嗾擬議佚催勧擬宥揃⇧泌爾試軌易籍葦磨爾淡、爾試某叢殆試晒議軌易爾淡(mitogine activated-protein kinase, MAPK)賜聾飲翌佚催距准爾淡(extracelluar signal-regulated kinases 1/2, ERK1/2) 吉吉。P70宰軌易悶爾淡(p70 ribosomal S6 kinase, p70S6爾淡)狼某葦磨/釦葦磨軌易爾淡社怛撹埀。輝聾飲歌嚥斤伏海咀徨、蛍叢吉壼豚郡哘扮⇧p70S6爾淡瓜爾試。猟孽烏祇壓白漬鱒彬議寄報麼強琢才伉縫伉壜⇧p70S6爾淡瓜儻堀爾試[1,5]。P70S6K宥狛距准mRNA議鍬咎狛殻糞孖斤聾飲伏海議距陣[6,7], p70S6爾淡議試晒鞭凪貧嗄宋擻次桓-3爾淡(phosphatidylinositol 3-kinse, PI3K)議距准[8]。猟孽[9,10]烏祇AngÅ 辛缶爾凍劍議VSMC才伉次聾飲 p70S6爾淡試來, 隼遇凪佚催勧擬余抄旺音賠萱⇧容霞嚥Ras-Raf-MEK-ERK1/2嗤購。恬宀參念議垢恬𠮟幣Raf 壓AngÅ 爾試ERK1/2狛殻嶄旺音軟麼勣恬喘⇧遇喇PKC-ζ函遇旗岻[11,12]。Saward 吉[13]烏祇1 μmol AngÅ嚥VSMC恬喘15 min辛爾試PI3K⇧聞PI3K p85冉汽了籍葦磨宋磨晒旺窟伏弔廬了。除定音富猟孽𠮟幣PKC-ζ頁PI3K議和嗄丼哘軌易, 揖扮匆歌嚥p70S6爾淡議距准, 凪試來鞭PI3K旗仍恢麗PtdIns(3-5)P(3)議距陣[14-16]。咀遇⇧厘断容霞PKC-ζ呀辛嬬初擬AngÅ爾試p70S6爾淡, 旺戻竃PI3K-PKC-ζ選栽距陣p70S6爾淡試來議垢恬邪譜。
, 為調匳勞
1 可創嚥圭隈
1.1 可創
森H-Ras森悶才森PKC-ζ森悶蛍艶杭徭Boehringer Mannheim才Santa Cruz巷望⇧森p70S6爾淡森悶才森p85PI3K森悶杭徭Upstate Biotechnology Inc巷望⇧S6訥喇紗鎮寄Kinetek Pharmaceuticals Inc巷望奘僕⇧[r-32P]ATP葎Dupont巷望恢瞳⇧Protein A-agarose 葎Life Technologies Inc巷望恢瞳⇧wortmannin、chelerythrine才rapamycin 葎Sigma巷望恢瞳⇧PKC-ζ邪來久麗杭徭Calbiochem巷望(La Jolla, CA)。
1.2 僮砿峠錆次聾飲凍劍
歌孚猟孽[17]⇧貫200゛250 g俛來Sprague Dawley寄報俟麼強琢蛍宣竃VSMC⇧喘根10%弌釘僮賠議DMEM凍劍児凍劍。函5゛13旗VSMC噐100 cm凍劍駐嶄凍劍崛70%゛80%蛮栽⇧喘根0.1%弌釘僮賠議DMEM凍劍児写偬凍劍48 h聞VSMC侃噐床峭彜蓑⇧隼朔喘AngÅ、 PDGF吉缶爾20 min。PI3K議雙崙質Wortmnnin、掲蒙呟來PKC-雙崙質Chelerythine、PKC-ζ 雙崙質(PKC-ζ邪來久麗)Pseudo Z才p70S6爾淡雙崙質rapamycin噐AngÅ、PDGF岻念10 min紗秘。
, http://www.100md.com
1.3 聾飲軌易戻函嚥敵業霞協
僮砿諸嫖殆Å(AngÅ)、 PDGF恬喘朔議VSMC喘宋磨産喝匣牢眉肝⇧耽駐紗1.0 mL TME産喝匣(10 mmol Tris, 5 mmol MgCl2, 1 mmol EDTA, 25 mmol NaF, pH 7.5◉匝喘念紗100 μmol Na3VO4, 20 mg/L leupeptin, 1 mg/L pepstatin A, 4 mg/L aprotinin, 1 mmol DTT) 叢盾聾飲。堀恭, 甥圈貧産蛸鹸釦⇧肯和聾飲⇧甥圈貧喘階蕗頚哩聾飲⇧15 000g宣伉30 min, 函貧賠匣⇧梓Bradford軌易霞協歳傍苧慕霞協軌易敵業⇧劔瞳廈贋噐-80≧棋喘。
1.4 p70S6爾淡試來霞協
歌孚猟孽[18]⇧函森p70S6爾淡森悶窒吽柿牛麗(50% bead slurry) 5 μL, 紗30 μL郡哘産喝匣(25 mmol b-glycerophosphate, 20 mmol MOPS, pH 7.2, 5 mmol EGTA, 2 mmol EDTA, 20 mmol MgCl2, 1 mmol Na3VO4, 0.25 mmol dithiothreitol), 紗4 μg 久麗S6膏訥才50 μmol [r-32 P]ATP(2〜1015 dpm/mol), 噐30≧郡哘15 min, 函20 μL郡哘詞栽麗泣劔噐2 cm2 p81 phosphocellulose 郡哘崕(Whatman巷望)⇧喘根1% orthophosphoric acid 肇宣徨邦喝牢眉肝⇧喘r-柴方匂芝村慧符試來。
, http://www.100md.com
1.5 Western Blot 糞刮
歌孚猟孽[11]⇧塘崙9%議SDS-PAGE蛍宣什才3%議持蚊什⇧耽迅紗秘20 μg軌易劔瞳⇧崔窮咯産喝匣嶄⇧70 V窮咯埃20 min。棋劔瞳序秘蛍宣什朔⇧150 V窮咯崛侭俶扮寂。 繍軌易廬咫崛嵓磨亘略院弔⇧喘Life Technologies巷望斌喘撃液匣噐片梁侃尖2 h⇧梓1|200紗p70S6爾淡、p85PI3K賜PKC-ζ汽森, 4≧狛匚⇧TBS匣牢弔10 min〜3肝⇧喘星功狛剳晒麗淡編質歳(ECL, Amersham International plc. United Kingdom)𠮟弼。
1.6 Ras-宋擻次桓-3爾淡-軌易爾淡C-ζ詞栽柿牛糞刮
歌孚猟孽[12]⇧函TME産喝匣戻函議軌易⇧紗森Ras汽森賜p85PI3K汽森才protein A-agarose噐4≧詞栽12 h序佩窒吽柿牛(immunoprecipitation, IP)。 Protein A-agarose喘TME産喝匣牢匯肝⇧TTBS産喝匣(20 mmol Tris, pH 7.5, 500 mmol NaCl, 1% Triton X-100, 0.1% beta-mercaptoethanol)牢屈肝⇧ 壅喘TME産喝匣牢匯肝。隼朔⇧喘9%議SDS-PAGE什繍窒吽鹸栽麗蛍宣⇧ 喘森p85PI3K才PKC-ζ汽森序佩窒吽咫治(immunoblot, IB)糞刮。
, 為調匳勞
1.7 聾飲柴方嚥覦炎俟寘獨爐用剳宰榠塊秘糞刮
歌孚猟孽[13]⇧函伏海措挫議VSMC崙撹聾飲傅匣(4〜107倖/L)俊嶽噐24迅凍劍医嶄⇧耽迅0.5 mL(2〜104倖聾飲)⇧剣圄24 h。算涙僮賠凍劍児写偬剣圄24 h朔⇧聞VSMC侃噐床峭豚⇧紗弌釘僮賠崛敵業葎10%⇧揖扮蛍怏紗秘100 nmol AngÅ(嶮敵業,和揖) 賜10 μg/L PDGF(嶮敵業,和揖)。 Wortmannin (嶮敵業10 nmol,和揖)、chelerythrine (嶮敵業10 μmol,和揖)才rapamycin(嶮敵業100 nmol,和揖)噐AngÅ、PDGF念10 min 紗秘⇧斤孚怏葎峪根10%弌釘僮賠議DMEM凍劍児, 写偬凍劍24 h。隼朔序佩和双糞刮: 〙聾飲嶧晒匣蛍宣聾飲⇧參4%岬杜清半弼柴方。〖 写偬凍劍18 h, 紗覦炎俟寘獨爐用剳宰榠(3 H-thymine deoxyribonucleotide, 3 H-TdR)崛嶮敵業葎2 mCi/L, 6 h朔辺劔⇧虹根3 H-TdR凍劍児⇧喘D-hanks匣牢眉肝⇧參聾飲嶧晒匣嶧晒。喘寔腎叡繍聾飲渇簾噐横詮亘略陀崕貧⇧5%眉柁閑磨耕協⇧涙邦厰桓用邦⇧孤夬朔崔描妨匿嶄紗描妨匣(遮噂500 mL, PPO 3, POPOP 0.1 g)噐匣悶描妨蛍裂卩(Packard 巷望 )霞慧符來膿業⇧參耽蛍嶝縫受方(decay per minute, dpm賜d/min)燕幣。
, http://www.100md.com
1.8 由柴侃尖
方象參
\s燕幣⇧方象将圭餓蛍裂朔序佩q殊刮⇧餓呟議𠮟广來喘P<0.05才P<0.01登僅。2 潤 惚
2.1 僮砿峠錆次聾飲燕器p70宰軌易悶S6爾淡、p85宋擻次桓-3爾淡才軌易爾淡C-ζ
Western Blot 蛍裂窟孖屎械凍劍議VSMC燕器p70S6爾淡、p85PI3K冉汽了才PKC-ζ, AngÅ才 PDGF缶爾旺音唹峒p70S6爾淡、p85PI3K冉汽了才PKC-ζ議燕器(夕1, Figure 1)。
夕1 僮砿峠錆次聾飲燕器p70宰軌易悶S6爾淡、p85宋擻次桓-3爾淡冉侏才軌易爾淡C-ζ
, http://www.100md.com
Figure 1 VSMC express p70S6K, p85PI3K subtype and PKC-ζ Western blot analysis was carried out with anti- p70S6K antibody (A), anti-p85PI3K antibody (B) and anti- p70S6K, antibody (C), respectively.
2.2 僮砿諸嫖殆Å爾試僮砿峠錆次聾飲p70宰軌易悶S6爾淡
夕2 (Figure 2) 𠮟幣AngÅ嚥VSMC恬喘5 min軸蝕兵爾試p70S6爾淡, 20 min器互桁⇧埃葎侃尖念議5蔚⇧昧朔幟愁週詰(A), 旺格質楚卆正來(B)。p70S6爾淡雙崙質rapamycin (100 nmol)、PI3K雙崙質wortmannin(10 nmol)才PKC-ζ雙崙質Pseudo Z (50 μmol)譲辛嗤丼怦僅AngÅ議恬喘(夕3, Figure 3)。
, 為調匳勞
2.3 僮砿諸嫖殆Å缶爾Ras-宋擻次桓-3爾淡-軌易爾淡C-ζ鹸栽麗侘撹
聾飲鞭AngÅ才 PDGF缶爾朔⇧辛鉱賀欺PI3K﨑Ras廬了旺嚥岻潤栽議孖嵆, 壓森Ras森悶柿牛議軌易嶄嗤PI3K贋壓。夕3A(Figure 3A)嶄辛需森Ras森悶柿牛議軌易窮咯蛍宣廬弔朔壅喘森PI3K森悶墫住辛𠮟幣p85訳揮⇧ wortmannin 10 nmol辛怦僅Ras-PI3K屢札潤栽。夕4B才夕4C(Figure 4B and 4C)嶄辛需音徽森Ras森悶辛繍PKC-ζ詞栽柿牛(B); 遇拝森PI3K森悶匆辛繍PKC-ζ柿牛和栖, PI3K、PKC-ζ岻寂議屢札恬喘呀辛瓜wortmannin才pseudo Z怦僅(C)。喇緩⇧厘断辛參容僅聾飲鞭AngÅ才 PDGF缶爾朔, Ras、PI3K、PKC-ζ眉嶽軌易潤栽壓匯軟侘撹匯倖孔嬬鹸栽悶。Wortmannin才pseudo Z辛怦僅宸倖孔嬬鹸栽悶議侘撹, 傍苧PI3K-PKC-ζ壓宸倖孔嬬鹸栽悶嶄軟嶷勣恬喘。
, 為調匳勞
夕2 僮砿諸嫖殆Å斤僮砿峠錆次聾飲p70宰軌易悶S6爾淡試來議唹峒
Figure 2 The effects of AngÅ on p70S6K activity of VSMC A⦿ Time-course of AngÅ on p70S6K activity. B⦿ Concentration-response of AngÅ on p70S6K activity.
夕3 p70宰軌易悶S6爾淡雙崙質rapamycin、宋擻次桓-3爾淡雙崙質wortmannin才軌易爾淡C-ζ雙崙質pseudo Z斤僮砿諸嫖殆Å試晒僮砿峠錆次聾飲p70宰軌易悶S6爾淡議唹峒
Figure 3 The effects of p70S6K inhibitor rapamycin⇧PI3K inhibitor wortmannin and PKC-ζ inhibitor pseudo Z on AngÅ activation of p70S6K in VSMC a⦿ P<0.01 compared with control grpip, b: P<0.01 compared with AngÅ group, c: P<0.01 compared with PDGF group.
, 為調匳勞
夕4 p85宋擻次桓-3爾淡才軌易爾淡C嚥Ras議選狼
Figure 4 Ras association with p85PI3K and PKC-ζ A and B. p85PI3K and PKC-ζ were immuno-precipitated (IP) with Ras antibody. C. PKC-ζ was immuno-precipitated (IP) by p85PI3K antibody.
2.4 Wortmannin才chelerythrine怦僅僮砿諸嫖殆Å嗾擬僮砿峠錆次聾飲奐岾
岬杜清半弼聾飲柴方才3 H-TdR塊秘隈蛍裂𠮟幣AngÅ(100 nmol)才PDGF (10 μg/L)譲嗤熟膿議缶爾VSMC奐岾議恬喘⇧AngÅ才PDGF侃尖怏聾飲柴方苧𠮟互噐斤孚怏⇧3 H-TdR塊秘楚𠮟广奐紗。Wortmannin 才chelerythrine 譲辛怦僅AngÅ才PDGF議恬喘(燕1, Table 1)。
, http://www.100md.com
燕1 僮砿諸嫖殆Å、僮弌医坿伏海咀徨、Wortmannin才chelerythrine斤僮砿峠錆次聾飲奐岾議唹峒
Table 1 Effects of Ang Å⇧ PDGF⇧ wortmannin and chelerythrine on VSMC proliferation (n=6,
\s) GroupsCell number
(〜104)
3 H-TdR incorporation
(dpm)
Control
, 為調匳勞
2.26\0.14
503\51
AngÅ
3.32\0.29a
740\75a
PDGF
3.51\0.46a
754\95a
Wortmannin
2.25\0.21
512\41
, 為調匳勞
AngÅ+Wortmannin
2.32\0.12b
540\34b
PDGF+Wortmannin
2.31\0.23c
542\60c
Chelerythrine
2.42\0.43
493\75
AngÅ+Chelerythrine
, 為調匳勞
2.47\0.43b
550\84b
PDGF+Chelerythrine
2.42\0.36c
560\71c
a:P<0.01 vs control; b: P<0.01 vs Ang Å◉ c: P<0.01 vs PDGF
3 網 胎
云猟議麼勣窟孖頁PKC-ζ才PI3K選栽距准AngÅ爾試僮砿峠錆次聾飲宰軌易悶爾淡p70S6爾淡。嗤泌和曾泣屶隔宸匯潤胎:〙AngÅ嚥VSMC恬喘5 min 軸蝕兵爾試p70S6爾淡, 20 min器互桁, 旺格質楚卆正來。P70S6K雙崙質rapamycin、 PI3K雙崙質wortmannin才PKC-ζ雙崙質(PKC邪來久麗) Pseudo Z譲辛苧𠮟怦僅AngÅ嗾擬議p70S6爾淡爾試◉ 〖 聾飲鞭AngÅ才 PDGF缶爾朔⇧侘撹Ras-PI3K-PKC-ζ孔嬬鹸栽悶⇧wortmannin才chelerythrine辛雙崙宸倖孔嬬鹸栽悶議侘撹。參念範葎AngÅ麼勣哈軟VSMC景寄, 除定窟孖AngÅ匆醤嗤伏海咀徨議蒙來⇧辛哈軟聾飲奐岾[1,13]。厮岑伏海咀徨宥狛凪聾飲弔貧議軌易籍葦磨鞭悶⇧繍佚催勧公Ras, Ras窟伏弔廬了旺爾試某葦磨/釦葦磨爾淡Raf, 朔宀頁軌易籍葦磨爾淡鞭悶佚催勧擬議嶄伉⇧Raf宥狛MEK爾試ERK1/2序佩佚催選雫慧寄⇧ERK1/2 (嗽各某叢殆爾試議爾淡, mitogen-activated protein kinases, MAPK)繍佚催勧崛聾飲宰⇧尼強DNA栽撹才屢購軌易栽撹, 陥序VSMC奐岾。隼遇⇧AngÅ 旺音恬喘噐軌易籍葦磨鞭悶, 遇頁恬喘噐7-TM鞭悶(seven transmembrane receptors, 7-TM)。7-TM鞭悶頁匯嶽G-軌易鞭悶, 旺音醤嗤籍葦磨爾淡試來。椎担⇧AngÅ頁奕劔陥序聾飲奐岾議椿? 音富猟孽烏祇戻幣軌易籍葦磨鞭悶嚥7-TM鞭悶岻寂贋壓屢札"斤三"(cross talk)[19゛21]。AngÅ辛処祇伏海咀徨佚催勧弓宥揃。猟孽[21⇧22]烏祇AngÅ辛爾試聾飲坪謹嶽軌易爾淡式凪佚催宥祇⇧泌Raf、PKC、PI3K、PLC吉吉。匯違範葎AngÅ辛宥狛曾訳余抄爾試p70S6爾淡: 匯訳辛嬬宥狛Ras-Raf-MEK-ERK1/2余抄。恬宀參念議冩梢窟孖Raf壓AngÅ爾試ERK1/2嶄旺音軟麼勣恬喘⇧遇喇PKC-ζ 函遇旗岻[11,12]。総匯訳辛嬬宥狛Ras-PI3K余抄[8゛10,12]⇧隼遇⇧凪峋聾宥揃旺音賠萱。 猟孽[14゛16]烏祇PKC-ζ歌嚥p70S6爾淡議距准旺初擬凪貧嗄PI3K議丼哘⇧PKC-ζ議試來卆正噐PI3K議恢麗PIP3。厘断容霞PKC-ζ才PI3K選栽歌嚥AngÅ爾試p70S6爾淡議距准。云糞刮鉱賀欺VSMC單米燕器p85PI3K才PKC-ζ, AngÅ質楚卆正來仇爾試p70S6爾淡。 Rapamycin、wortmannin才Pseudo Z譲辛怦僅AngÅ議恬喘。嗤箸議頁⇧厘断窟孖聾飲鞭AngÅ才 PDGF缶爾朔⇧侘撹Ras-PI3K-PKC-ζ孔嬬鹸栽悶⇧森Ras森悶才森PI3K森悶辛詞栽柿牛PKC-ζ, wortmannin才pseudo Z譲辛怦僅緩鹸栽悶議侘撹。緩翌⇧厘断喘VSMC奐岾恬葎嶮泣峺炎蛍裂阻PI3K才PKC-ζ議恬喘⇧ 鉱賀欺PI3K雙崙質wortmannin才掲蒙呟來PKC雙崙質chelerythrine譲辛苧𠮟雙崙AngÅ嗾擬議VSMC奐岾。潤栽參念議垢恬⇧厘断戻竃AngÅ爾試僮砿峠錆次聾飲p70S6爾淡辛嬬淫根泌和佚催勧擬余抄⦿AngÅ 恬喘噐VSMC AT1鞭悶⇧爾試PI3K, 試晒議PI3K嗾擬PKC-ζ匯揖嚥Ras潤栽侘撹孔嬬鹸栽悶, 旺爾試PKC-ζ, 恷嶮爾試p70S6爾淡, 擬崑VSMC奐岾(夕5, Figure 5)。
, 為調匳勞
夕5 僮砿諸嫖殆Å将喇Ras-宋淡次桓-3爾淡-軌易爾淡C-ζ爾試p70宰軌易悶S6爾淡議余抄
Figure 5 AngÅ activates p70S6K via the Ras-PI3K-PKC-ζ pathway.
[崑仍] 云猟何蛍垢恬壓胆忽鯖腹禽寄僥Dr. Berk糞刮片才紗鎮寄悟戴曳冉寄僥Dr. Pelech糞刮片頼撹。
[児署𡸴朕] 忽社徭隼親僥児署彿廁仁籾(39970847)
[恬宀酒初] 僧極啓⇧槻⇧刷掴福邃臭偏繁⇧1959定竃伏⇧鴬平冩梢伏。購喟坿⇧槻⇧鴻叫福繁⇧嶄表匳親寄僥勞尖僥縮娩⇧鴬平冩梢伏擬弗⇧忽暦垪僥了溜埀氏勞僥親得咏怏撹埀⇧ゞ嶄忽勞尖僥宥烏〃械暦園溜⇧嶄忽勞尖僥氏伉僮砿廨匍溜埀氏械暦溜埀。
歌深猟孽
, http://www.100md.com
[1] Berk BC. Angiotensin Å signal transduction in vascular smooth muscle: pathways activated by specific tyrosine kinases [J]. J Am Soc Nephrol⇧ 1999⇧ 10: S62-S68
[2] Taubman MB, Berk BC, Izumo S, et al. Angiotensin Å induces c-fos mRNA in aortic smooth muscle cells: role of Ca2+ mobilization and protein kinase C activation [J]. J Biol Chem, 1989, 264: 526-530
[3] Molloy CJ, Taylor DS, Weber H. Angiotensin Å stimulation of rapid protein tyrosine phosphorylation and protein kinase activation in rat aortic smooth muscle cells [J]. J Biol Chem, 1993, 268 (10): 7 338-345
, 為調匳勞
[4] Duff JL, Berk BC, Corson MA. Angiotensin Å stimulates the pp44 and pp42 mitogen-activated protein kinases in cultured rat aortic smooth muscle cells [J]. Biochem Biophys Res Commun, 1992, 188: 257-264
[5] Griendling KK, Ushio-Fukai M, Lassegue B, et al. Angiotensin Å signaling in vascular smooth muscle. New concepts [J]. Hypertension, 1997, 29 (1 Pt 2): 366-373
[6] Pullen N, Thomas G. The modular phosphorylation and activation of p70S6k [J]. FEBS Lett, 1997, 410 (1): 78-82
, 為調匳勞
[7] Avruch J. Insulin signal transduction through protein kinase cascades [J]. Mol Cell Biochem, 1998, 182 (1-2): 31-48
[8] Chung J, Grammer TC, Lemon KP, et al. PDGF- and insulin-dependent pp70S6k activation mediated by phosphatidylinositol-3-OH kinase [J]. Nature, 1994, 370 (6484): 71-75
[9] Giasson E, Meloche S. Role of p70S6 protein kinase in angiotensin Å-induced protein synthesis in vascular smooth muscle cells [J]. J Biol Chem, 1995, 270 (10): 5 225-231
, 為調匳勞
[10] Sadoshima J, Izumo S. Rapamycin selectively inhibits angiotensin Å-induced increase in protein synthesis in cardiac myocytes in vitro. Potential role of 70-kD S6 kinase in angiotensin Å-induced cardiac hypertrophy [J]. Circ Res, 1995, 77 (6): 1 040-052
[11] Liao DF, Duff JL, Daum G, et al. Angiotension Å stimulates MAP kinase kinase kinase activity in vascular smooth muscle cell. Role of Raf [J]. Circ Res, 1996, 79 (5): 1 007-014
, 為調匳勞
[12] Liao DF, Monia B, Dean N, et al. Protein kinase C-ζ mediates angiotensin Å activation of ERK1/2 in vascular smooth muscle cells [J]. J Biol Chem, 1997, 272: 6 146-150
[13] Saward L, Zahradka P. Angiotensin Å activates phosphatidylinositol 3-kinase in vascular smooth muscle cells [J]. Circ Res, 1997, 81 (2): 249-257
[14] Chou MM, Hou W, Johnson J, et al. Regulation of protein kinase C zeta by PI3-kinase and PDK-1 [J]. Curr Biol, 1998, 8 (19): 1 069-077
, 為調匳勞
[15] Le Good JA, Ziegler WH, Parekh DB, et al. Protein kinase C isotypes controlled by phosphoinositide 3-kinase through the protein kinase PDK1 [J]. Science, 1998, 281 (5385): 2 042-045
[16] Romanelli A, Martin KA, Toker A, et al. p70S6 kinase is regulated by protein kinase C zeta and participates in a phosphoinositide 3-kinase-regulated signalling complex [J]. Mol Cell Biol, 1999, 19 (4): 2 921-928
[17] Liao DF, Jin ZG, Baas AS, et al. Purification and identification of secreted oxidative stress-induced factors from vascular smooth muscle cells [J]. J Biol Chem, 2000, 275 (1): 189-196
, 為調匳勞
[18] Xu YJ, Ouk Kim S, Liao DF, et al. Stimulation of 90- and 70-kDa ribosomal protein S6 kinases by arginine vasopressin and lysophosphatidic acid in rat cardiomyocytes [J]. Biochem Pharmacol, 2000, 59 (9): 1 163-171
[19] Velloso LA, Folli F, Sun XJ, et al. Cross-talk between the insulin and angiotensin signaling systems [J]. Proc Natl Acad Sci USA, 1996, 93 (22): 12 490-495
[20] Eguchi S, Numaguchi K, Iwasaki H, et al. Calcium-dependent epidermal growth factor receptor transactivation mediates the angiotensin Å-induced mitogen-activated protein kinase activation in vascular smooth muscle cells [J]. J Biol Chem, 1998, 273 (15): 8 890-896
, 為調匳勞
[21] Berk BC, Corson MA. Angiotensin Å signal transduction in vascular smooth muscle: role of tyrosine kinases [J]. Circ Res, 1997, 80 (5): 607-616
[22] Eguchi S, Iwasaki H, Ueno H, et al. Intracellular signaling of angiotensin Å-induced p70S6 kinase phosphorylation at Ser(411) in vascular smooth muscle cells. Possible requirement of epidermal growth factor receptor, Ras, extracellular signal-regulated kinase, and Akt [J]. J Biol Chem, 1999, 274 (52): 36 843-851
(緩猟2000-07-11辺欺, 2000-09-05俐指), http://www.100md.com