白细胞介素-4基因修饰Raji细胞促进肿瘤特异细胞毒性T细胞杀伤活性
作者:赵晓东 李成荣 杨锡强 黄贵清 付劲蓉 蒋莉萍 王莉佳
单位:400014 重庆医科大学儿童医院
关键词:白细胞介素-4;伯基特;淋巴瘤;基因;T淋巴细胞;细胞毒性
中华儿科杂志990309 【摘要】 目的 探讨人白细胞介素(IL)-4基因修饰对伯基特淋巴瘤细胞株――Raji的体外生物学特性、肿瘤特异性细胞毒性T细胞(CTL)杀伤活性的影响及其机制。方法 采用逆转录病毒介导基因转染法将人IL-4 cDNA导入Raji株并筛选获得高效价表达IL-4的克隆,观察IL-4表达克隆形态学特征、体外增殖反应及其诱导的外周血单个核细胞(PBMC)增殖反应和IL-2、干扰素(IFN-)γ基因表达及CTL杀伤活性。结果 IL-4基因修饰对Raji的体外生物学特性无明显影响,但明显促进正常人PBMC增殖及IL-2、IFN-γ基因表达,其诱导建立的CTL杀伤活性明显增强,效应细胞:靶细胞比例为30∶1时,IL-4基因修饰可使Raji细胞诱导的CTL杀伤活性上升2~3倍。结论 外源性IL-4基因表达产物可能作用于瘤细胞和(或)PBMC,通过诱导IL-2、IFN-γ等细胞因子促进 CTL反应,并增强其杀伤活性。
, http://www.100md.com
Stimulation with IL-4 gene modified Raji cells enhances killing activity of tumor-specific cytotoxic T lymphocyte ZHAO Xiaodong, LI Chengrong, YANG Xiqiang, et al. Children′s Hospital, Chongqing University of Medical Science, Chongqing 400014
【Abstract】 Objective Effects of interleukin-4 (IL-4) gene modification of biological characteristics of Raji cell line and killing activity of Raji-specific cytotoxic T lymphocyte (CTL) were investigated. Methods IL-4 cDNA was amplified from a plasmid, pCD-hIL-4, by polymerase chain reaction (PCR) and transferred into Raji cells via retrovirus-mediated gene transfection method. Proliferation of IL-4-producing Raji clones and PBMC, IL-2 and IFN-γ mRNA synthesis and killing activity of Raji cell-induced CTL were determined. Results Stimulation with IL-4 gene-modified Raji cell increased proliferation and IL-2, IFN-γ mRNA synthesis of normal PBMC, which could not be blocked by anti-IL-4 monoclonal antibody. The killing activity of CTL induced by IL-4 gene modified Raji clones was much stronger than that of controls. Conclusion IL-4 gene modification is capable of enhancing killing activity of Raji-specific CTL by means of up-regulating expression of IL-2 and creating a more suitable environment for antigen-presenting, establishment and maintenance of killing function of CTL.
, 百拇医药
【Key words】 Interleukin-4 Burkitt′s lymphoma Genes T-lymphocytes, cytotoxic
恶性淋巴瘤在儿科临床实践中并不少见,目前的治疗手段尚未取得令人满意的效果。采用细胞因子基因修饰的肿瘤细胞疫苗激发机体抗肿瘤免疫功能,最终彻底清除肿瘤是一种具有潜在临床价值的新方法。将其与手术切除、放疗、化疗结合进行治疗可望大大改善恶性淋巴瘤患儿的预后。本研究将白细胞介素(IL-4)基因导入伯基特(Burkitt)淋巴瘤株――Raji,观察其体外生物学特性的改变及对 T细胞功能的影响,旨在为恶性淋巴瘤的基因与免疫治疗提供新线索。
材料及方法
一、目的基因、载体及宿主菌
以含人IL-4 cDNA全序列的质粒PCD-hIL-4为模板,用引物Sense 5′-AACTCGAGTTAATGGGTCTCA CCTCCCAA-3′和Antisense 5′-AGGGATCCTATTCAG CTCGAACACTTTGA-3′通过聚合酶链反应(PCR)扩增获得IL-4 cDNA片段,琼脂糖凝胶电泳回收、纯化该片段。用双脱氧链终止法测序。逆转录病毒载体(PLXSN)由Miller(USA)赠送,长约 5 900碱基对。宿主菌大肠杆菌Mc 1 061株由作者单位的免疫室保存。
, http://www.100md.com
二、主要试剂、细胞株及其来源
内切酶、T4 DNA连接酶、Taq DNA聚合酶及逆转录酶均购自Promega公司;脱氧三磷酸核苷(dNTP)、Polybrene、细胞培养基RPMI 1 640、DMEM和克隆筛选试剂G 418购自Gibco公司;脂质体转染试剂Dosper及重组IL-4购自Boehringer Mannheim公司;51 铬酸钠(51Cr)购自美国Ameshan公司, 3 H-TdR购自中国同位素公司,IL-2、干扰素(IFN-)γ PCR引物购自上海细胞生物研究所;重组 IL-4(rIL-4)、鼠抗人IL-4单克隆抗体及IL-4酶联免疫吸附试验(ELISA)检测试剂盒购自 Genzyme公司。病毒包装细胞株PA317及病毒滴度鉴定细胞株 NIH3T3来自第三军医大学分子生物室, Raji为本室常规传代培养。
三、重组逆转录病毒载体的构建、包装、筛选及感染
, 百拇医药
人IL-4重组逆转录病毒载体构建策略如图1所示。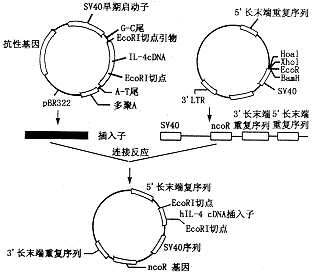
图1 IL-4重组逆转录病毒载体构建示意图
碱裂解法抽提质粒,酶切、连接、鉴定及细菌转化等步骤均按 Sambrook等[1]介绍的标准方法操作。具体步骤为:用平端内切酶 HpaI使载体线性化。酚:氯仿抽提纯化后与 PCR反应扩增回收得到的目的基因片段,在T4 DNA连接酶催化作用下进行连接反应。连接反应产物转化感受态大肠杆菌-Mc1 061,将其接种于琼脂平板,待菌落形成后进行大肠杆菌扩增,抽提质粒,酶切后琼脂糖电泳鉴定。脂质体介导基因转染、病毒滴度鉴定及感染 Raji细胞方法见参考文献[2]。
四、表达IL-4的Raji细胞克隆筛选及体外生物学特性观察
, http://www.100md.com
用G 418筛选转基因 Raji细胞克隆1个月,收集抗性细胞克隆上清液用ELISA法检测IL-4表达水平。在普通光学显微镜下观察抗性Raji克隆形态学特征,用3H-TdR掺入法测定其体外增殖活性。
五、Raji细胞诱导正常人外周血单个核细胞(PBMC)增殖及IL-2、IFN-γ基因表达
常规分离正常人PBMC,调细胞浓度至1×106/ml,加100 μl入55孔板,不同组别加入经丝裂霉素C(40 μg/L×106细胞,37℃,1小时)处理的转基因或对照 Raji细胞。 IL-4单克隆抗体(简称单抗)阻断组加入终浓度为0.5 μg/ml的鼠抗人 IL-4单抗,重组 IL-4组加入终浓度为0.1 μg/ml的IL-4,37℃、5%二氧化碳(CO2)培养24小时。培养结束前16小时加入3H-TdR,液闪计数每分钟脉冲数(cpm)值。另取1×106 PBMC加入24孔板,分别加入丝裂霉素C处理的肿瘤细胞1×105/孔(100 μl),置37℃、5%CO2孵箱中培养12、24、48和72小时后抽提总RNA,用RT-PCR检测IL-2和IFN-γ基因表达。
, http://www.100md.com
六、Raji特异性细胞毒性T淋巴细胞(CTL)建立及其杀伤活性测定
参照Thomas等[3]介绍的方法。用 51 Cr标记野生型肿瘤细胞(100 μci/1×106细胞,37℃过夜),PBS缓冲液洗涤3次后将浓度调至1×105/ml作为靶细胞。采用4小时 51 Cr释放法测定CTL杀伤活性。于96孔培养板中加入不同比例的效应细胞和靶细胞(每种比例重复3孔),37℃、5%CO2孵育4小时,1 500 r/min离心5分钟,收集上清用γ-计数仪测定其cpm值。按下式计算CTL杀伤活性,以杀伤率%表示:
%杀伤活性=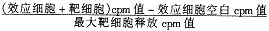 ×100
×100
结 果
, http://www.100md.com
一、重组IL-4逆转录病毒载体的构建
用核酸内切酶SacI消化载体出现2片段,消化重组体后2片段大小相近,无法区分;DNA内切酶EcoRⅠ消化重组体出现367 bp小片段,消化载体后无此片段出现,证实正向重组的 IL-4逆转录病毒构建成功(图2)。
图2 重组IL-4逆转录病毒载体酶切鉴定图谱
二、Raji株表达 IL-4水平及其体外生物学特征
各株Raji细胞克隆IL-4表达水平如表1所示。导入IL-4cDNA的Raji细胞产生IL-4的最高水平24小时可达134.5 pg/106细胞。普通光学显微镜下观察IL-4高表达克隆细胞形态、大小与野生型Raji及仅导入载体的Raji细胞(Raji-neo)相比无明显变化。其体外增殖反应水平与对照组比较亦无明显差异。
, http://www.100md.com
表1 Raji细胞表达IL-4水平及其体外增殖反应 细胞株
IL-4表达水平
(pg/106细胞24h)
增殖反应(cpm)
未加组
rIL-4组
野生型Raji
0
2 213
2 376
Raji-neo
, 百拇医药 0
2 437
2 643
R-IL-41
134.5
2 678
-
R-IL-42
87.4
2 734
-
R-IL-43
, 百拇医药
63.8
2 765
-
R-IL-44
21.7
1 988
-
注:-示未检测;每一实验至少重复3次;
R-IL-41~4示IL-4基因修饰的Raji株,下同
三、Raji细胞诱导的正常人PBMC增殖反应
如表2所示,表达IL-4的4株Raji克隆诱导的正常人PBMC增殖反应明显强于野生型Raji及Raji-neo诱导者,IL-4单抗不能阻断。重组人IL-4加入培养系统虽可使PBMC增殖反应增强,但其程度远不及表达IL-4的Raji克隆。
, 百拇医药
表2 Raji细胞诱导的正常人PBMC体外增殖反应(cpm) 诱导细胞株
未加组
IL-4单抗组
rIL-4组
空白对照
412
-
564
野生型Raji
619
-
1 386
, 百拇医药
Raji-neo
546
-
1 748
R-IL-41
3 865
3 647
-
R-IL-42
3 467
3 282
-
, 百拇医药
R-IL-43
3 180
3 254
-
R-IL-44
2 789
2 403
-
四、Raji细胞诱导正常人PBMC IL-2、IFN-γ基因表达
IL-4基因修饰的Raji细胞可诱导正常人PBMC IL-2和IFN-γ基因表达。IL-2基因表达在刺激后12小时即可检测,维持至48小时开始减弱;IFN-γ基因刺激后12小时有较弱表达,至24小时达表达高峰,48小时后开始减弱(图3、4)。野生型Raji、Raji-neo、加入rIL-4共培养的Raji及单独加入rIL-4均未能诱导上述基因表达。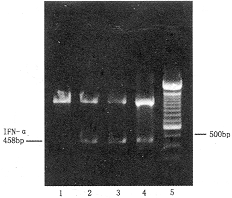
, 百拇医药
1:Raji-rIL-4;2:Raji-neo;3:R-IL-4,24h;
4:R-IL-4;5:Marker
图3 R-IL-41诱导的正常人PBMC IL-2基因表达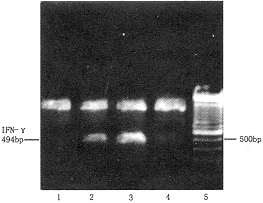
1:Raji+rIL-4;2:Raji-neo;3:R-IL-4,24h;
4:R-IL-41;12h;5:Marker
图4 R-Il-41诱导的正常人PBMC IFN-γ基因表达
五、CTL的杀伤活性
, http://www.100md.com
野生型Raji细胞和Raji-neo仅能诱导轻度CTL杀伤活性。各种效应细胞:靶细胞比例两者诱导的杀伤活性无显著差异。R-IL-41细胞诱导的CTL杀伤活性明显增强,并随效应细胞:靶细胞比例的升高而增强效应细胞∶靶细胞比例为30∶1时Raji-neo、野生型Raji及R-IL-41诱导的CTL杀伤效率分别为(20.3±6.2)%、(20.8±4.3)%和(48.8±10.4)%。讨 论
细胞因子与机体抗肿瘤免疫反应密切相关,已证实诸多细胞因子在肿瘤局部表达可激发机体抗肿瘤免疫反应,提示细胞因子基因修饰肿瘤疫苗具有潜在的临床应用前景。 IL-4是一种具有多种生物学功能的细胞因子,前期研究发现 IL-4基因修饰的小鼠实体瘤细胞可增强宿主的抗肿瘤免疫反应[4];重组 IL-4可明显抑制新鲜分离的非何杰金淋巴瘤细胞体外增殖反应[5],提示 IL-4既可能直接作用于肿瘤细胞,抑制其增殖或促进其细胞凋亡过程[6],亦可上调机体抗肿瘤免疫反应。
, http://www.100md.com
本研究发现 IL-4基因修饰 Raji细胞诱导的正常人PBMC增殖反应及 IL-2, IFN-γ基因表达明显增强,而加入培养系统的重组 IL-4却无此作用。提示由肿瘤细胞自分泌的 IL-4可能更有效地作用于瘤细胞本身和(或) PBMC,最终促进PBMC增殖并表达一些有利于抗原提呈、 CTL反应建立及 CTL杀伤活性的细胞因子产生,从而有利于机体有效清除肿瘤。IL-2能促进 T细胞增殖,诱导 T细胞分泌一系列细胞因子,亦为CD8+CTL维持杀伤活性必需的生长因子,故推测其可能为 IL-4基因修饰瘤苗促进 CTL杀伤活性的关键作用环节。
IL-4主要来源于TH2细胞,具有诱导T细胞向TH2方向分化并产生TH2类细胞因子的功能。本研究发现IL-4基因修饰瘤苗主要诱导正常人PBMC产生 IL-2、 IFN-γ等TH1细胞类细胞因子, 而用rIL-4或rIL-4加肿瘤细胞则未能诱导PBMC产生上述细胞因子, 提示肿瘤细胞分泌的 IL-4可能并未直接作用于 T细胞。我们推测IL-4基因修饰 Raji细胞所致的继发因素,如肿瘤细胞膜表面某些抗原的改变, 可能激活 T淋巴细胞受体, 从而诱导建立TH1应答。 CTL是抗肿瘤免疫网络中最特异、最重要的效应细胞,本研究结果为 IL-4基因修饰淋巴瘤疫苗的使用提供了线索,但尚需在体内进一步证实。
, 百拇医药
本课题为国家自然科学基金资助(编号:39470733)及卫生部科研课题基金资助(编号:96-1-366)
参考文献
1 Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: a laboratory manual. New York: Cold Spring Harbor Laboratory Press, 1989. 27-110.
2 赵晓东,李成荣,杨锡强,等.人IL-4重组逆转录载体的构建及在人肿瘤细胞中的表达.重庆医科大学学报,1998,1:1-4.
3 Thomas F, Gajewski, Francesca F, et al. Tumor rejection requires a CTLA4 ligant provided by the host or expressed on the tumor. J Immunol, 1996, 156:2909-2922.
, 百拇医药
4 曹雪涛,于益芝,徐志工.IL-4基因转染瘤苗体内抗肿瘤转移作用及共免疫机理的初步研究.中华微生物学和免疫学杂志,1995,15:53-55.
5 Defrance T, Fluckiger AC, Rossi JF, et al. Antiproliferative effects of interleukin-4 on freshli isolated non-Hodgkin malignant B-lymphoma cells. Blood, 1992, 79:990-996.
6 Manabe A, Coustan Smith E, Kumagai M, et al. Interleukin-4 induces programmed cell death (Apoptosis) in cases of high-risk acute lymphoblastic leukemia. Blood, 1994, 83:1731-1737.
(收稿:1998-02-26 修回:1998-07-24), 百拇医药
单位:400014 重庆医科大学儿童医院
关键词:白细胞介素-4;伯基特;淋巴瘤;基因;T淋巴细胞;细胞毒性
中华儿科杂志990309 【摘要】 目的 探讨人白细胞介素(IL)-4基因修饰对伯基特淋巴瘤细胞株――Raji的体外生物学特性、肿瘤特异性细胞毒性T细胞(CTL)杀伤活性的影响及其机制。方法 采用逆转录病毒介导基因转染法将人IL-4 cDNA导入Raji株并筛选获得高效价表达IL-4的克隆,观察IL-4表达克隆形态学特征、体外增殖反应及其诱导的外周血单个核细胞(PBMC)增殖反应和IL-2、干扰素(IFN-)γ基因表达及CTL杀伤活性。结果 IL-4基因修饰对Raji的体外生物学特性无明显影响,但明显促进正常人PBMC增殖及IL-2、IFN-γ基因表达,其诱导建立的CTL杀伤活性明显增强,效应细胞:靶细胞比例为30∶1时,IL-4基因修饰可使Raji细胞诱导的CTL杀伤活性上升2~3倍。结论 外源性IL-4基因表达产物可能作用于瘤细胞和(或)PBMC,通过诱导IL-2、IFN-γ等细胞因子促进 CTL反应,并增强其杀伤活性。
, http://www.100md.com
Stimulation with IL-4 gene modified Raji cells enhances killing activity of tumor-specific cytotoxic T lymphocyte ZHAO Xiaodong, LI Chengrong, YANG Xiqiang, et al. Children′s Hospital, Chongqing University of Medical Science, Chongqing 400014
【Abstract】 Objective Effects of interleukin-4 (IL-4) gene modification of biological characteristics of Raji cell line and killing activity of Raji-specific cytotoxic T lymphocyte (CTL) were investigated. Methods IL-4 cDNA was amplified from a plasmid, pCD-hIL-4, by polymerase chain reaction (PCR) and transferred into Raji cells via retrovirus-mediated gene transfection method. Proliferation of IL-4-producing Raji clones and PBMC, IL-2 and IFN-γ mRNA synthesis and killing activity of Raji cell-induced CTL were determined. Results Stimulation with IL-4 gene-modified Raji cell increased proliferation and IL-2, IFN-γ mRNA synthesis of normal PBMC, which could not be blocked by anti-IL-4 monoclonal antibody. The killing activity of CTL induced by IL-4 gene modified Raji clones was much stronger than that of controls. Conclusion IL-4 gene modification is capable of enhancing killing activity of Raji-specific CTL by means of up-regulating expression of IL-2 and creating a more suitable environment for antigen-presenting, establishment and maintenance of killing function of CTL.
, 百拇医药
【Key words】 Interleukin-4 Burkitt′s lymphoma Genes T-lymphocytes, cytotoxic
恶性淋巴瘤在儿科临床实践中并不少见,目前的治疗手段尚未取得令人满意的效果。采用细胞因子基因修饰的肿瘤细胞疫苗激发机体抗肿瘤免疫功能,最终彻底清除肿瘤是一种具有潜在临床价值的新方法。将其与手术切除、放疗、化疗结合进行治疗可望大大改善恶性淋巴瘤患儿的预后。本研究将白细胞介素(IL-4)基因导入伯基特(Burkitt)淋巴瘤株――Raji,观察其体外生物学特性的改变及对 T细胞功能的影响,旨在为恶性淋巴瘤的基因与免疫治疗提供新线索。
材料及方法
一、目的基因、载体及宿主菌
以含人IL-4 cDNA全序列的质粒PCD-hIL-4为模板,用引物Sense 5′-AACTCGAGTTAATGGGTCTCA CCTCCCAA-3′和Antisense 5′-AGGGATCCTATTCAG CTCGAACACTTTGA-3′通过聚合酶链反应(PCR)扩增获得IL-4 cDNA片段,琼脂糖凝胶电泳回收、纯化该片段。用双脱氧链终止法测序。逆转录病毒载体(PLXSN)由Miller(USA)赠送,长约 5 900碱基对。宿主菌大肠杆菌Mc 1 061株由作者单位的免疫室保存。
, http://www.100md.com
二、主要试剂、细胞株及其来源
内切酶、T4 DNA连接酶、Taq DNA聚合酶及逆转录酶均购自Promega公司;脱氧三磷酸核苷(dNTP)、Polybrene、细胞培养基RPMI 1 640、DMEM和克隆筛选试剂G 418购自Gibco公司;脂质体转染试剂Dosper及重组IL-4购自Boehringer Mannheim公司;51 铬酸钠(51Cr)购自美国Ameshan公司, 3 H-TdR购自中国同位素公司,IL-2、干扰素(IFN-)γ PCR引物购自上海细胞生物研究所;重组 IL-4(rIL-4)、鼠抗人IL-4单克隆抗体及IL-4酶联免疫吸附试验(ELISA)检测试剂盒购自 Genzyme公司。病毒包装细胞株PA317及病毒滴度鉴定细胞株 NIH3T3来自第三军医大学分子生物室, Raji为本室常规传代培养。
三、重组逆转录病毒载体的构建、包装、筛选及感染
, 百拇医药
人IL-4重组逆转录病毒载体构建策略如图1所示。
图1 IL-4重组逆转录病毒载体构建示意图
碱裂解法抽提质粒,酶切、连接、鉴定及细菌转化等步骤均按 Sambrook等[1]介绍的标准方法操作。具体步骤为:用平端内切酶 HpaI使载体线性化。酚:氯仿抽提纯化后与 PCR反应扩增回收得到的目的基因片段,在T4 DNA连接酶催化作用下进行连接反应。连接反应产物转化感受态大肠杆菌-Mc1 061,将其接种于琼脂平板,待菌落形成后进行大肠杆菌扩增,抽提质粒,酶切后琼脂糖电泳鉴定。脂质体介导基因转染、病毒滴度鉴定及感染 Raji细胞方法见参考文献[2]。
四、表达IL-4的Raji细胞克隆筛选及体外生物学特性观察
, http://www.100md.com
用G 418筛选转基因 Raji细胞克隆1个月,收集抗性细胞克隆上清液用ELISA法检测IL-4表达水平。在普通光学显微镜下观察抗性Raji克隆形态学特征,用3H-TdR掺入法测定其体外增殖活性。
五、Raji细胞诱导正常人外周血单个核细胞(PBMC)增殖及IL-2、IFN-γ基因表达
常规分离正常人PBMC,调细胞浓度至1×106/ml,加100 μl入55孔板,不同组别加入经丝裂霉素C(40 μg/L×106细胞,37℃,1小时)处理的转基因或对照 Raji细胞。 IL-4单克隆抗体(简称单抗)阻断组加入终浓度为0.5 μg/ml的鼠抗人 IL-4单抗,重组 IL-4组加入终浓度为0.1 μg/ml的IL-4,37℃、5%二氧化碳(CO2)培养24小时。培养结束前16小时加入3H-TdR,液闪计数每分钟脉冲数(cpm)值。另取1×106 PBMC加入24孔板,分别加入丝裂霉素C处理的肿瘤细胞1×105/孔(100 μl),置37℃、5%CO2孵箱中培养12、24、48和72小时后抽提总RNA,用RT-PCR检测IL-2和IFN-γ基因表达。
, http://www.100md.com
六、Raji特异性细胞毒性T淋巴细胞(CTL)建立及其杀伤活性测定
参照Thomas等[3]介绍的方法。用 51 Cr标记野生型肿瘤细胞(100 μci/1×106细胞,37℃过夜),PBS缓冲液洗涤3次后将浓度调至1×105/ml作为靶细胞。采用4小时 51 Cr释放法测定CTL杀伤活性。于96孔培养板中加入不同比例的效应细胞和靶细胞(每种比例重复3孔),37℃、5%CO2孵育4小时,1 500 r/min离心5分钟,收集上清用γ-计数仪测定其cpm值。按下式计算CTL杀伤活性,以杀伤率%表示:
%杀伤活性=
×100结 果
, http://www.100md.com
一、重组IL-4逆转录病毒载体的构建
用核酸内切酶SacI消化载体出现2片段,消化重组体后2片段大小相近,无法区分;DNA内切酶EcoRⅠ消化重组体出现367 bp小片段,消化载体后无此片段出现,证实正向重组的 IL-4逆转录病毒构建成功(图2)。
图2 重组IL-4逆转录病毒载体酶切鉴定图谱
二、Raji株表达 IL-4水平及其体外生物学特征
各株Raji细胞克隆IL-4表达水平如表1所示。导入IL-4cDNA的Raji细胞产生IL-4的最高水平24小时可达134.5 pg/106细胞。普通光学显微镜下观察IL-4高表达克隆细胞形态、大小与野生型Raji及仅导入载体的Raji细胞(Raji-neo)相比无明显变化。其体外增殖反应水平与对照组比较亦无明显差异。
, http://www.100md.com
表1 Raji细胞表达IL-4水平及其体外增殖反应 细胞株
IL-4表达水平
(pg/106细胞24h)
增殖反应(cpm)
未加组
rIL-4组
野生型Raji
0
2 213
2 376
Raji-neo
, 百拇医药 0
2 437
2 643
R-IL-41
134.5
2 678
-
R-IL-42
87.4
2 734
-
R-IL-43
, 百拇医药
63.8
2 765
-
R-IL-44
21.7
1 988
-
注:-示未检测;每一实验至少重复3次;
R-IL-41~4示IL-4基因修饰的Raji株,下同
三、Raji细胞诱导的正常人PBMC增殖反应
如表2所示,表达IL-4的4株Raji克隆诱导的正常人PBMC增殖反应明显强于野生型Raji及Raji-neo诱导者,IL-4单抗不能阻断。重组人IL-4加入培养系统虽可使PBMC增殖反应增强,但其程度远不及表达IL-4的Raji克隆。
, 百拇医药
表2 Raji细胞诱导的正常人PBMC体外增殖反应(cpm) 诱导细胞株
未加组
IL-4单抗组
rIL-4组
空白对照
412
-
564
野生型Raji
619
-
1 386
, 百拇医药
Raji-neo
546
-
1 748
R-IL-41
3 865
3 647
-
R-IL-42
3 467
3 282
-
, 百拇医药
R-IL-43
3 180
3 254
-
R-IL-44
2 789
2 403
-
四、Raji细胞诱导正常人PBMC IL-2、IFN-γ基因表达
IL-4基因修饰的Raji细胞可诱导正常人PBMC IL-2和IFN-γ基因表达。IL-2基因表达在刺激后12小时即可检测,维持至48小时开始减弱;IFN-γ基因刺激后12小时有较弱表达,至24小时达表达高峰,48小时后开始减弱(图3、4)。野生型Raji、Raji-neo、加入rIL-4共培养的Raji及单独加入rIL-4均未能诱导上述基因表达。
, 百拇医药
1:Raji-rIL-4;2:Raji-neo;3:R-IL-4,24h;
4:R-IL-4;5:Marker
图3 R-IL-41诱导的正常人PBMC IL-2基因表达
1:Raji+rIL-4;2:Raji-neo;3:R-IL-4,24h;
4:R-IL-41;12h;5:Marker
图4 R-Il-41诱导的正常人PBMC IFN-γ基因表达
五、CTL的杀伤活性
, http://www.100md.com
野生型Raji细胞和Raji-neo仅能诱导轻度CTL杀伤活性。各种效应细胞:靶细胞比例两者诱导的杀伤活性无显著差异。R-IL-41细胞诱导的CTL杀伤活性明显增强,并随效应细胞:靶细胞比例的升高而增强效应细胞∶靶细胞比例为30∶1时Raji-neo、野生型Raji及R-IL-41诱导的CTL杀伤效率分别为(20.3±6.2)%、(20.8±4.3)%和(48.8±10.4)%。讨 论
细胞因子与机体抗肿瘤免疫反应密切相关,已证实诸多细胞因子在肿瘤局部表达可激发机体抗肿瘤免疫反应,提示细胞因子基因修饰肿瘤疫苗具有潜在的临床应用前景。 IL-4是一种具有多种生物学功能的细胞因子,前期研究发现 IL-4基因修饰的小鼠实体瘤细胞可增强宿主的抗肿瘤免疫反应[4];重组 IL-4可明显抑制新鲜分离的非何杰金淋巴瘤细胞体外增殖反应[5],提示 IL-4既可能直接作用于肿瘤细胞,抑制其增殖或促进其细胞凋亡过程[6],亦可上调机体抗肿瘤免疫反应。
, http://www.100md.com
本研究发现 IL-4基因修饰 Raji细胞诱导的正常人PBMC增殖反应及 IL-2, IFN-γ基因表达明显增强,而加入培养系统的重组 IL-4却无此作用。提示由肿瘤细胞自分泌的 IL-4可能更有效地作用于瘤细胞本身和(或) PBMC,最终促进PBMC增殖并表达一些有利于抗原提呈、 CTL反应建立及 CTL杀伤活性的细胞因子产生,从而有利于机体有效清除肿瘤。IL-2能促进 T细胞增殖,诱导 T细胞分泌一系列细胞因子,亦为CD8+CTL维持杀伤活性必需的生长因子,故推测其可能为 IL-4基因修饰瘤苗促进 CTL杀伤活性的关键作用环节。
IL-4主要来源于TH2细胞,具有诱导T细胞向TH2方向分化并产生TH2类细胞因子的功能。本研究发现IL-4基因修饰瘤苗主要诱导正常人PBMC产生 IL-2、 IFN-γ等TH1细胞类细胞因子, 而用rIL-4或rIL-4加肿瘤细胞则未能诱导PBMC产生上述细胞因子, 提示肿瘤细胞分泌的 IL-4可能并未直接作用于 T细胞。我们推测IL-4基因修饰 Raji细胞所致的继发因素,如肿瘤细胞膜表面某些抗原的改变, 可能激活 T淋巴细胞受体, 从而诱导建立TH1应答。 CTL是抗肿瘤免疫网络中最特异、最重要的效应细胞,本研究结果为 IL-4基因修饰淋巴瘤疫苗的使用提供了线索,但尚需在体内进一步证实。
, 百拇医药
本课题为国家自然科学基金资助(编号:39470733)及卫生部科研课题基金资助(编号:96-1-366)
参考文献
1 Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: a laboratory manual. New York: Cold Spring Harbor Laboratory Press, 1989. 27-110.
2 赵晓东,李成荣,杨锡强,等.人IL-4重组逆转录载体的构建及在人肿瘤细胞中的表达.重庆医科大学学报,1998,1:1-4.
3 Thomas F, Gajewski, Francesca F, et al. Tumor rejection requires a CTLA4 ligant provided by the host or expressed on the tumor. J Immunol, 1996, 156:2909-2922.
, 百拇医药
4 曹雪涛,于益芝,徐志工.IL-4基因转染瘤苗体内抗肿瘤转移作用及共免疫机理的初步研究.中华微生物学和免疫学杂志,1995,15:53-55.
5 Defrance T, Fluckiger AC, Rossi JF, et al. Antiproliferative effects of interleukin-4 on freshli isolated non-Hodgkin malignant B-lymphoma cells. Blood, 1992, 79:990-996.
6 Manabe A, Coustan Smith E, Kumagai M, et al. Interleukin-4 induces programmed cell death (Apoptosis) in cases of high-risk acute lymphoblastic leukemia. Blood, 1994, 83:1731-1737.
(收稿:1998-02-26 修回:1998-07-24), 百拇医药