基因多态性对情绪调节神经回路的影响(2)
 |
Arialysis),,即评估一个脑区活动的程度与另一个脑区活动的相关程度,Harid等人(2000)确认了腹外侧前额叶对杏仁核的调节关系。用类似范式开展的神经心理学研究表明,前额叶与杏仁核的连接障碍可能是造成情绪障碍患者行为抑制能力差的主要原因(Altshuler,Bookheimer,Townsend et al.,2005;Blumberg,Leung,Skudlarski et al.,2003)。
1.3皮层与边缘系统的相互作用
在总结动物情绪和人类情绪脑成像研究的基础上,Ochsner等人(2004;2007)以自下而上和自上而下的观点构建了情绪调节的神经结构模型(见图1)。该模型认为情绪调节涉及不同层次的多种评价系统的交互作用,大脑神经网络一方面用自下而上的方式编码刺激的情绪属性,对不同类型情绪进行评价加工,从而产生不同的情绪反应,另一方面也以自上而下的方式执行对情绪刺激的评价和情绪表达或体验的控制。在结构上,自下而上的情绪产生系统处在皮层后部和皮层下区域,其主要结构包括基底核和杏仁核等边缘系统;而自上而下的情绪控制系统位于大脑的皮层区域,主要是额叶区,且包含有两种不同类型的系统,第一类系统是自上而下的描述性评价系统(Description-Based Appraisal System,DBAS),包括背侧前额叶和扣带区,负责对刺激的联系、情绪的特征和状态等产生心理描述:另一类系统是自上而下以结果为基础的评价系统(Outcome-Based Appraisal System,OBAS),包括眶前额叶、腹侧前额叶和扣带区,负责表征情绪反应的选择与结果之间的联系(Ochsner&Gross,2007)。DBAS和OBAS一起参与对皮层下情绪反应执行不同类型的控制。DBAS运用高水平认知功能来调节情绪,相反,OBAS则通过被动的条件性和工具性学习获得的行为模式对情绪进行调节,但是这两个系统的不同成分如何对不同类型情绪进行不同的控制还不清楚,这些系统如何与其它系统相互作用,如何参与非情绪的“冷”控制,如何在积极的调节控制中发挥作用等问题还有待进一步的研究。
, http://www.100md.com
归纳起来,背侧前额叶和腹侧前额叶连接,前额叶和杏仁核连接,皮层,边缘系统连接构成了情绪调节的主要神经回路,不同神经回路在情绪调节的神经网络系统中起着不同的作用,其他的一些直接和间接的神经网络,网络中具体节点的内部关系也对情绪调节产生重要的影响(Taylor,Liberzon,2007),如果情绪调节神经回路的结构和功能出现障碍将容易导致情绪疾病的产生。这些研究成果使得我们对情绪调节的神经基础有了新的认识,然而要弄清情绪控制信息在这些回路中的运行过程,参与情绪调节的神经回路之间是如何相互作用的,还需进一步从细胞、基因等微观层次上理解情绪调节的神经生物学作用机制。
2 基因多态性对情绪调节神经回路的影响
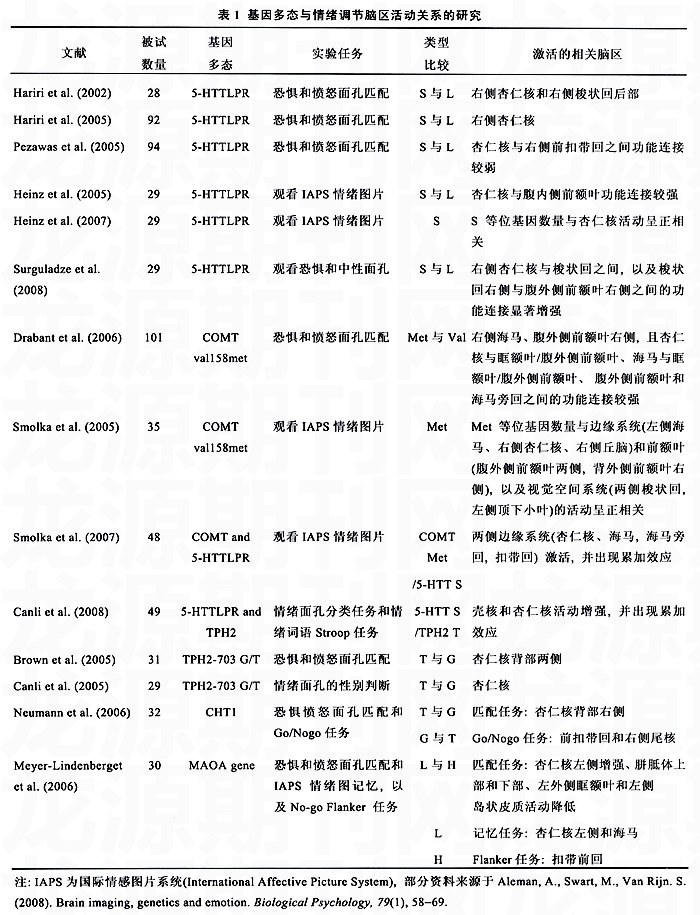
近年来,人们对基因多态性或基因变异在大脑结构和功能变化中所起的作用越来越感兴趣。基因变异导致特定神经结构的发展出现遗传差异,而这些差异会影响神经元的信息传递,最终使得这些脑区更有可能出现异常,因此,基因的功能多态性在一定程度上调控着特定行为的结果。在传统的基因与行为关联研究的基础上,随着后基因时代认知神经科学的发展,一种新近发展迅速的基因影像学研究方法在“基因与脑”和“脑与行为”之间架起一座“看得见的桥梁”,使得人们可以在脑结构层次上以更少的被试,更客观的测量手段理解基因对行为的影响。基因影像学(Imaging Genetics)运用生理影像技术作为表型分析来评价基因的变异情况,探讨基因是如何影响大脑的神经结构和功能,以及由此导致的心理病理(Hariri,Drabant&Weinberger,2006)。正是以此为研究方法,不同领域研究者考察了不同基因多态对情绪加工脑区及其神经信息传递的影响,使得情绪调节神经回路的研究获得了突破性进展。
, 百拇医药
2.1 5-羟色胺传运体
大脑中一种名为5-羟色胺(5-hydroxytrypta-mine,5-HT)的化学物质在调节人的情绪方面有着重要的作用,5-羟色胺的活动像其他单胺类神经递质一样,是由细胞质膜里的5-羟色胺转运体(5-hydroxytryptamine transporter,5-HTT)来调节的。随着5-羟色胺的释放,位于突触前的5-羟色胺转运体把5-羟色胺返回细胞用于再循环或降低代谢率。在正常的生理条件下,5-羟色胺运转体的重摄取是清除细胞外5-羟色胺的主要手段,它通过调节突触后受体介导信号的作用量和作用时间,调控着情绪加工脑区神经元信息传递的正常活动(canli&Lesch,2007)。因此,5-羟色胺转运体是情绪调节的重要的候选基因,获得了较多研究关注(见表1)。
人类5-HTT基因(SLC6A4)位于17q11.1-12染色体上,在它的控制转录的启动部位即5-HTT基因一连锁多态区(5-HTT-linked polymorphic region,5-HTTLPR)的上游大约1kb处,含有一个44bp的插入或缺失突变,从而形成L型(528bp)和s型(484bp)两种等位基因,构成三种基因型L/L、L/S和S/S。s型等位基因表现低转录活性,携带S型等位基因的5-HTT回吸收能力低,而L型等位基因表现高转录活性,与5-HTT快速回收有关(Lesch,Bengel,Heils et al.,1996)。研究表明s等位基因携带者与心境障碍、抑郁和焦虑症等情绪疾病有较强的正相关(Stein,Westenberg,Liebowitz,2002)。其主要表现为s等位基因携带者杏仁核的反应活动出现异常兴奋。这说明s等位基因携带者负责情绪的神经活动受到影响,使其对负性情绪更加敏感,为情绪疾病提供了潜在的神经基础。
杏仁核是情绪加工的核心脑区,也是5-羟色胺密集部位,如果5-羟色胺转运体基因出现遗传差异,将导致杏仁核活动出现异常。例如,Pezawas等人(2005)运用基于体素的形态学研究方法(voxel-based morphometry,VBM)发现。相比L等位基因,s等位基因携带者参与情绪调节的前额叶和扣带回膝部fthe perigenual anterior cingulate codex,pAcc)的灰质容积显著的减小了。当给被试呈现生气和恐惧面孔时,与L/L等位纯合子相比,S/S或L/S等位基因携带者的扣带回膝部脑区的活动更低,而且s等位基因携带者的前扣带回和杏仁核的功能连接相对较弱,他们认为前额叶皮层和杏仁核结构连接强度的减弱影响了它们在功能上的相互作用,导致前额叶皮, 百拇医药(陈桃林 罗跃嘉)