5-羟色胺转运体基因多态性与强迫症的相关性研究(2)
 |
 |
1.2.3电泳分型(1)5-HTTVNTR电泳分型。12% PAGE 180 V电泳90 min,溴化乙啶(EB)染色20 min,拍照分型。267 bp为等位基因10,301 bp为等位基因12。(2)5-HTTLPR电泳分型。3%琼脂糖凝胶电泳100 V电泳100 min,EB染色2O min,拍照分型。367 bp为等位基因S,411 bp为等位基因L。
1.3统计学方法
全部数据输入计算机,使用SPSS13.0统计软件进行数据分析。方法包括频数分析、Hardy-Weinberg平衡法则(H-W定律)的吻合度检验、χ2检验等。
2结果
2.1OCD组和对照组的H-W定律吻合度检验
OCD组观察值与期望值吻合良好(χ2=2.71,v=1,P>0.05);对照组观察值和期望值吻合良好(χ2=2.86,v=1,P>0.05);均符合H-W定律。
, http://www.100md.com
2.2OCD组与对照组间SLC6A4基因型及等位基因频数分布的比较
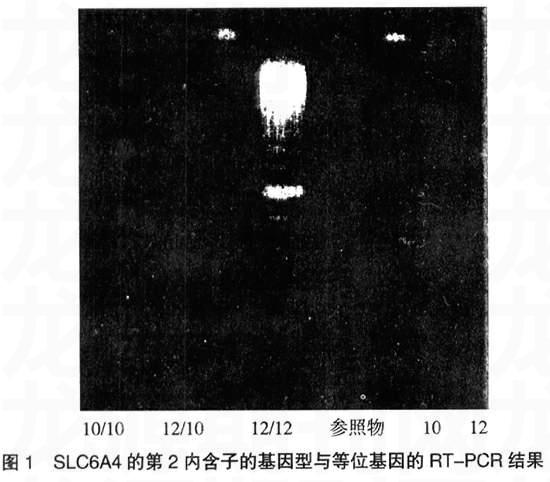
见表1,由表1可见,OCD组与对照组的SLC6A4第2内含子基因型(χ2=10.42,P<0.05)和等位基因总体(χ2=7.53,P<0.01)分布的差异有显著性或非常显著性,OCD组第2内含子基因型及等位基因PCR图见图1;两组的SLC6A4启动子的基因型分布的差异有显著性(χ2=6.061,P<0.05)。进一步比较两组的同性别间基因型和等位基因分布,差异无显著性。
2.3OCD组及对照组不同性别间的SLC6A4基因型分布比较
见表2。OCD组内不同性别间SLC6A4启动子(χ2=2.64,P>0.05)和第2内含子(χ2=0.50,P>0.05)的基因型分布、对照组内不同性别间SLC6A4启动子(χ2=2.64,P>0.05)和第2内含子(χ2=1.87,P>0.05)基因型分布的差异均无显著性。
, 百拇医药
2.4SLC6A4基因型与OCD的关联分析
关联分析结果显示,OCD患者与5-HITVNTR等位基因l0[比数比(OR)=2.24,Z=2.69,P<0.05,95%可信区间(95%CI)=1.24 ~4.02]及12/10基因型(OR=2.12,Z=2.13,P<0.05,95%CI=1.06 ~4.13)呈正关联,与5-HTTLPR基因L/L基因型亦呈正关联(OR=3.57,Z=2.39,P<0.05,95%CI=1.26~10.15)。在根据发病年龄、Y-BOCS评分等进行分组后比较,未发现该基因的基因型或等位基因分布的差异有显著性。
3讨论
五羟色胺(5-HT)是体内重要的神经递质,有多种突触前后受体亚型,且能被五羟色胺转运体(5-HTT)这种单一蛋白转运入突触前神经元。5-HTT从突触间隙中移除5-HT而决定突触后受体介导信号的量和作用持续时间,从而在5-HT神经传递的微调中起关键作用。5-HTT是一种约为630个氨基酸残基的蛋白质,在不同种属及不同细胞的克隆中,氨基酸残基数目有微小差异。人和鼠的5-HTT基因分别位于7号和11号染色体上,两者均由跨度约为35kb的14个外显子组成,有保守的外显子/内含子结构及相对较少的5’端非编码区和调节序列。人类5-HTT促进子区有一个多态性重复序列(5-HTTLPR),位于其转录起始位点上游约lkb处。大部分等位基因由14(short,S)或16(1ong,L)个22bp的重复序列组成,两种等位基因的频率在不同人种中存在差异,亚洲人种中S型基因频率明显高于高加索人种,且存在罕见的18或20个重复序列。该多态性对5-HTT 的转录有调节作用,LL型纯合子比SS型或LS型的原始淋巴细胞细胞株产生更高浓度的5-HTTmRNA。另有研究表明,尸检标本中LS、SS型个体脑干缝核5-HTTmRNA 含量显著低于LL型个体。在5-HTT基因中还有一个位于内含子2上的17bp的可变数目串联重复序列(VNTR)多态性,包括1O(STin2.10)、12(STin2.12)个重复的常见序列和9(STin2.9)个重复的罕见序列。由于上述两种多态性可能影响5-HTT基因的表达,许多研究者进行了一系列与情感障碍、精神分裂症、物质依赖、人格特征、强迫症等疾病的相关研究,试图从基因多态性来发现这些疾病的易感因素。
, 百拇医药
强迫症(OCD)以持续的强迫性思维和仪式性动作为特征,家族和双生子研究提示其与遗传因素有关。大多数用于治疗强迫症的药都具有抑制5-HT回收的作用,从突触末端回收5-HT是通过5-HTT 中介的,药物作用于5-HTT,可能是导致强迫症症状改善的一系列神经化学反应的关键。McDougle等[4]在34个欧裔美国家庭三代人中检验5-HTTLPR等位基因和OCD的连锁不平衡,35个杂合子父母中24个传递“L”型等位基因,11个传递“S”等位基因,如果单独分析13个五羟色胺重摄取抑制剂无反应的患者,其中1O个传递了“L”基因,3个传递了“S”基因,作者认为5-HTTLPR“L”基因和OCD可能有连锁不平衡。而Ramoz等[3]通过涉及352个家庭的大型队列研究了传递“L”或”S”等位基因5-HTTLPR基因对个人的影响,此外,筛选5个先前公布的与OCD相关的单核苷酸多态性(SNPs)的5'区域SLC6A4以及另外4个单核苷酸多态性5'区域。结论认为SLC6A4变种似乎没有参与OCD。Camarena等[5]对墨西哥人群的研究也不支持5-HTTLPR等位基因L与OCD的关系及未发现前述的连锁不平衡。Denys等[6]报道在OCD患者中未发现5-HTTVNTR基因型或等位基因分布与正常对照的差异有显著性。Baca-Garcia等[7]报道5-HTTVNTR的多态性等位基因与OCD的发生有关。
, 百拇医药
本结果显示,SLC6A4基因同样存在第2内含子VNTR多态和启动子Ins/Del多态,同时还发现5-HTTVNTR的基因型分布和等位基因频率在OCD和健康对照之间的差异均有显著性,OCD患者5-HTTLPR的基因型分布与健康对照之间的差异也有显著性。OCD与5-HTTVNTR等位基因l0(低重复次数)和12/10基因型呈正关联,携带等位基因10和基因型为12/10的人群发展为OCD的危险性分别是无等位基因10和非12/10基因型人群的2.24倍和2.12倍。OCD与5-HTTLPR基因型L/L呈正关联,基因型为L/L的人群发展为OCD的危险性可能是非L/L表型人群的3.57倍。但未发现这种差异有性别间的不同,不支持男女OCD患者可能具有不同发病机制的观点。国外报道5-HTTVNTR等位基因l2与等位基因l0相比,可以增加报告基因的表达水平;5-HTTLPR等位基因S与等位基因L相比,可限制SLC6A4基因调控区的转录,从而导致SLC6A4基因的低效表达[2,8-9]。因此这两个多态位点不同的等位基因和基因型会引起SLC6A4基因表达水平的变化,造成5-HT转运过程异常,进一步引起体内5-HT含量或分布部位的变化,参与了OCD的发病过程。然而,5-HT系统在其代谢和执行生物功能的过程中还有许多重要结构,本结果不能解释全部的临床现象。同时,OCD是一种病因复杂的多基因遗传疾病,可能在多种易感性基因特殊组合的情况下才会导致该病的发生,因此SLC6A4基因的基因型12/10和L/L可能只是增加了OCD发生的可能性。同时,OCD的临床表现千变万化,不同的临床亚型可能有不同的发病机制,而心理、社会和环境因素在OCD的发病中可能也具有一定作用。因此,在进一步的研究中需寻找更敏感的OCD亚型分类指标,应尽可能减少样本的异质性,并探讨环境与基因的相互作用,以期找到OCD的遗传病理机制。, 百拇医药(冯冬梅 甘露春 梁颂游 刘金英 代 娟 石 乔 韩自力 陶 炯)